

Cutting edge: stage-specific requirement of IL-18 for antiviral NK cell expansion
- PMID: 25589075
- PMCID: PMC4323636
- DOI: 10.4049/jimmunol.1402001
Cutting edge: stage-specific requirement of IL-18 for antiviral NK cell expansion
Abstract
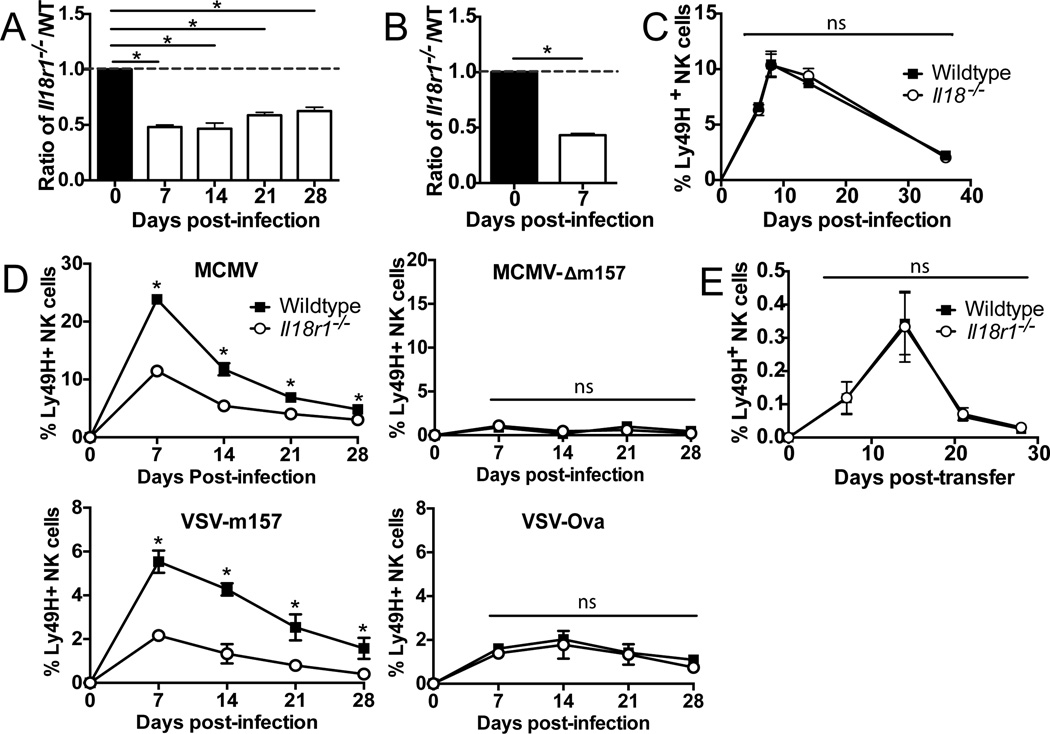
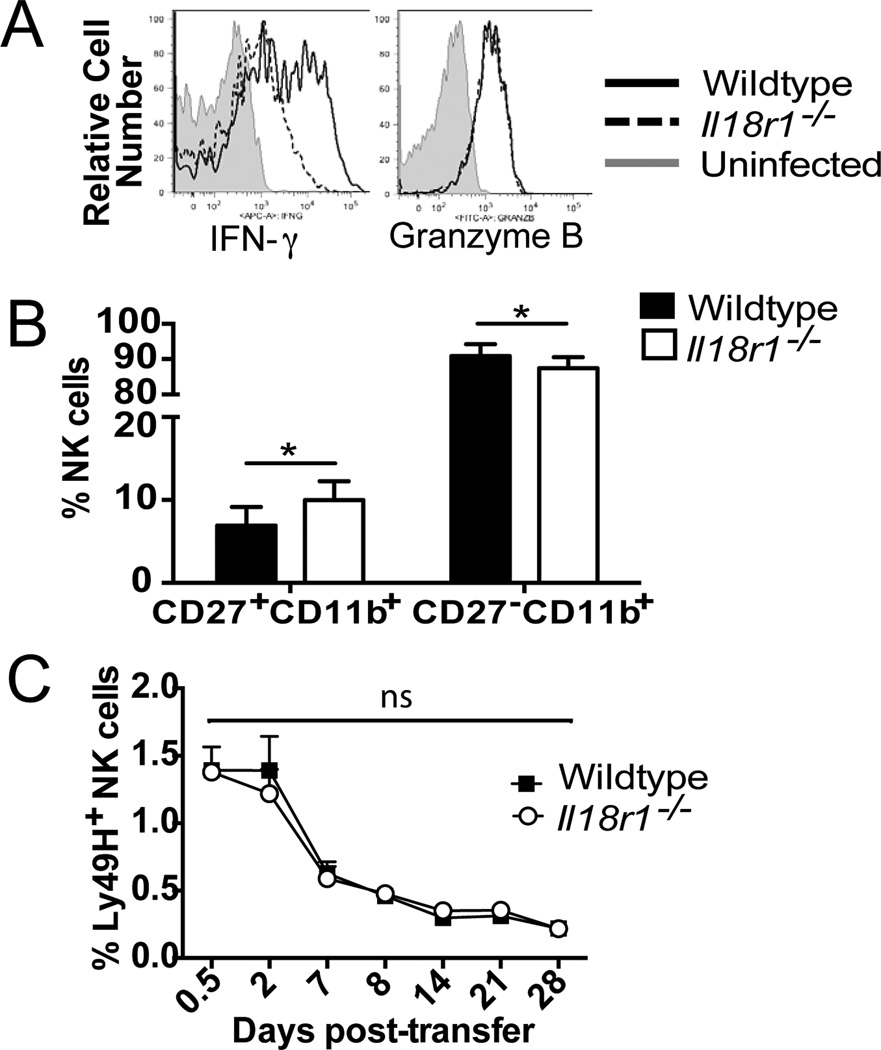
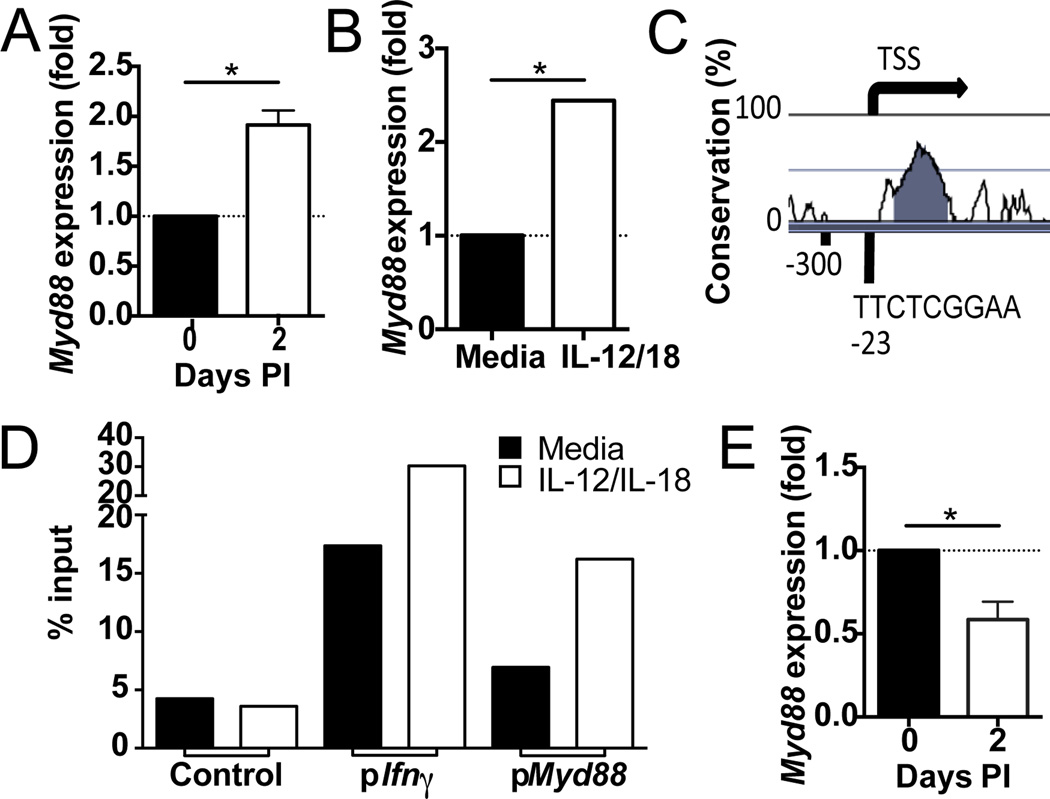
Although NK cells are considered part of the innate immune system, recent studies have demonstrated the ability of Ag-experienced NK cells to become long-lived and contribute to potent recall responses similar to T and B cells. The precise signals that promote the generation of a long-lived NK cell response are largely undefined. In this article, we demonstrate that NK cells require IL-18 signaling to generate a robust primary response during mouse CMV (MCMV) infection but do not require this signal for memory cell maintenance or recall responses. IL-12 signaling and STAT4 in activated NK cells increased the expression of the adaptor protein MyD88, which mediates signaling downstream of the IL-18 and IL-1 receptors. During MCMV infection, NK cells required MyD88, but not IL-1R, for optimal expansion. Thus, an IL-18-MyD88 signaling axis facilitates the prolific expansion of NK cells in response to primary viral infection, but not recall responses.
Copyright © 2015 by The American Association of Immunologists, Inc.
Conflict of interest statement
The authors declare no financial conflicts of interest.
Figures


Similar articles
-
Molecular Programming of Immunological Memory in Natural Killer Cells.Adv Exp Med Biol. 2015;850:81-91. doi: 10.1007/978-3-319-15774-0_7. Adv Exp Med Biol. 2015. PMID: 26324348 Review.
-
Proinflammatory cytokine signaling required for the generation of natural killer cell memory.J Exp Med. 2012 May 7;209(5):947-54. doi: 10.1084/jem.20111760. Epub 2012 Apr 9. J Exp Med. 2012. PMID: 22493516 Free PMC article.
-
Cytomegalovirus generates long-lived antigen-specific NK cells with diminished bystander activation to heterologous infection.J Exp Med. 2014 Dec 15;211(13):2669-80. doi: 10.1084/jem.20141172. Epub 2014 Nov 24. J Exp Med. 2014. PMID: 25422494 Free PMC article.
-
Stage-specific regulation of natural killer cell homeostasis and response against viral infection by microRNA-155.Proc Natl Acad Sci U S A. 2013 Apr 23;110(17):6967-72. doi: 10.1073/pnas.1304410110. Epub 2013 Apr 9. Proc Natl Acad Sci U S A. 2013. PMID: 23572582 Free PMC article.
-
Immunoregulatory cytokine networks: 60 years of learning from murine cytomegalovirus.Med Microbiol Immunol. 2015 Jun;204(3):345-54. doi: 10.1007/s00430-015-0412-3. Epub 2015 Apr 8. Med Microbiol Immunol. 2015. PMID: 25850988 Free PMC article. Review.
Cited by
-
Dendritic cells and natural killer cells: The road to a successful oncolytic virotherapy.Front Immunol. 2023 Jan 10;13:950079. doi: 10.3389/fimmu.2022.950079. eCollection 2022. Front Immunol. 2023. PMID: 36703982 Free PMC article. Review.
-
Transcription factor ID2 prevents E proteins from enforcing a naïve T lymphocyte gene program during NK cell development.Sci Immunol. 2018 Apr 27;3(22):eaao2139. doi: 10.1126/sciimmunol.aao2139. Sci Immunol. 2018. PMID: 29703840 Free PMC article.
-
The Multifaceted Roles of NK Cells in the Context of Murine Cytomegalovirus and Lymphocytic Choriomeningitis Virus Infections.Immune Netw. 2024 Jun 27;24(4):e29. doi: 10.4110/in.2024.24.e29. eCollection 2024 Aug. Immune Netw. 2024. PMID: 39246620 Free PMC article. Review.
-
Control of nutrient uptake by IRF4 orchestrates innate immune memory.Nat Immunol. 2023 Oct;24(10):1685-1697. doi: 10.1038/s41590-023-01620-z. Epub 2023 Sep 11. Nat Immunol. 2023. PMID: 37697097 Free PMC article.
-
Cutting Edge: STAT4 Promotes Bhlhe40 Induction to Drive Protective IFN-γ from NK Cells during Viral Infection.J Immunol. 2023 Nov 15;211(10):1469-1474. doi: 10.4049/jimmunol.2300402. J Immunol. 2023. PMID: 37830760 Free PMC article.
References
-
- Biron CA, Nguyen KB, Pien GC, Cousens LP, Salazar-Mather TP. Natural killer cells in antiviral defense: function and regulation by innate cytokines. Annu Rev Immunol. 1999;17:189–220. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
