

Absence of the uracil DNA glycosylase of murine gammaherpesvirus 68 impairs replication and delays the establishment of latency in vivo
- PMID: 25589640
- PMCID: PMC4337548
- DOI: 10.1128/JVI.03111-14
Absence of the uracil DNA glycosylase of murine gammaherpesvirus 68 impairs replication and delays the establishment of latency in vivo
Abstract
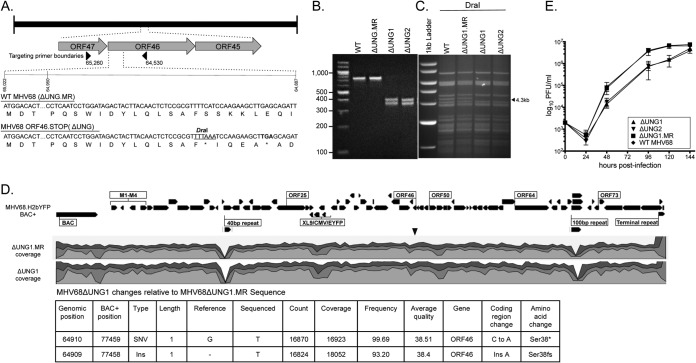
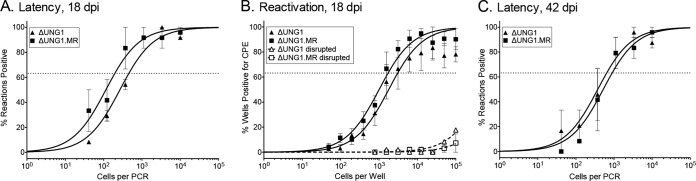
Uracil DNA glycosylases (UNG) are highly conserved proteins that preserve DNA fidelity by catalyzing the removal of mutagenic uracils. All herpesviruses encode a viral UNG (vUNG), and yet the role of the vUNG in a pathogenic course of gammaherpesvirus infection is not known. First, we demonstrated that the vUNG of murine gammaherpesvirus 68 (MHV68) retains the enzymatic function of host UNG in an in vitro class switch recombination assay. Next, we generated a recombinant MHV68 with a stop codon in ORF46/UNG (ΔUNG) that led to loss of UNG activity in infected cells and a replication defect in primary fibroblasts. Acute replication of MHV68ΔUNG in the lungs of infected mice was reduced 100-fold and was accompanied by a substantial delay in the establishment of splenic latency. Latency was largely, yet not fully, restored by an increase in virus inoculum or by altering the route of infection. MHV68 reactivation from latent splenocytes was not altered in the absence of the vUNG. A survey of host UNG activity in cells and tissues targeted by MHV68 indicated that the lung tissue has a lower level of enzymatic UNG activity than the spleen. Taken together, these results indicate that the vUNG plays a critical role in the replication of MHV68 in tissues with limited host UNG activity and this vUNG-dependent expansion, in turn, influences the kinetics of latency establishment in distal reservoirs.
Importance: Herpesviruses establish chronic lifelong infections using a strategy of replicative expansion, dissemination to latent reservoirs, and subsequent reactivation for transmission and spread. We examined the role of the viral uracil DNA glycosylase, a protein conserved among all herpesviruses, in replication and latency of murine gammaherpesvirus 68. We report that the viral UNG of this murine pathogen retains catalytic activity and influences replication in culture. The viral UNG was impaired for productive replication in the lung. This defect in expansion at the initial site of acute replication was associated with a substantial delay of latency establishment in the spleen. The levels of host UNG were substantially lower in the lung compared to the spleen, suggesting that herpesviruses encode a viral UNG to compensate for reduced host enzyme levels in some cell types and tissues. These data suggest that intervention at the site of initial replicative expansion can delay the establishment of latency, a hallmark of chronic herpesvirus infection.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures








References
-
- Nilsen H, Steinsbekk KS, Otterlei M, Slupphaug G, Aas PA, Krokan HE. 2000. Analysis of uracil-DNA glycosylases from the murine Ung gene reveals differential expression in tissues and in embryonic development and a subcellular sorting pattern that differs from the human homologues. Nucleic Acids Res 28:2277–2285. - PMC - PubMed
-
- Kavli B, Sundheim O, Akbari M, Otterlei M, Nilsen H, Skorpen F, Aas PA, Hagen L, Krokan HE, Slupphaug G. 2002. hUNG2 is the major repair enzyme for removal of uracil from U:A matches, U:G mismatches, and U in single-stranded DNA, with hSMUG1 as a broad specificity backup. J Biol Chem 277:39926–39936. doi:10.1074/jbc.M207107200. - DOI - PubMed
-
- Akbari M, Otterlei M, Pena-Diaz J, Aas PA, Kavli B, Liabakk NB, Hagen L, Imai K, Durandy A, Slupphaug G, Krokan HE. 2004. Repair of U/G and U/A in DNA by UNG2-associated repair complexes takes place predominantly by short-patch repair both in proliferating and growth-arrested cells. Nucleic Acids Res 32:5486–5498. doi:10.1093/nar/gkh872. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
