

DAMP signaling is a key pathway inducing immune modulation after brain injury
- PMID: 25589753
- PMCID: PMC4293412
- DOI: 10.1523/JNEUROSCI.2439-14.2015
DAMP signaling is a key pathway inducing immune modulation after brain injury
Erratum in
-
Erratum: Liesz et al., "DAMP Signaling Is a Key Pathway Inducing Immune Modulation after Brain Injury".J Neurosci. 2019 Jul 3;39(27):5419. doi: 10.1523/JNEUROSCI.0878-19.2019. Epub 2019 Jun 14. J Neurosci. 2019. PMID: 31201233 Free PMC article. No abstract available.
Abstract
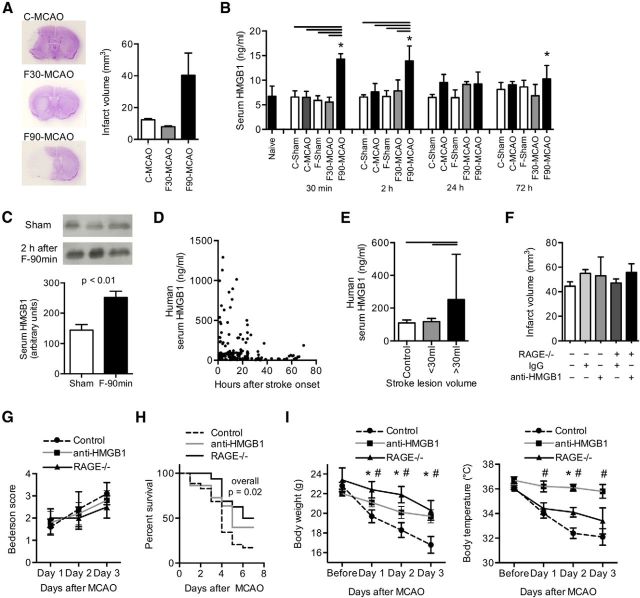
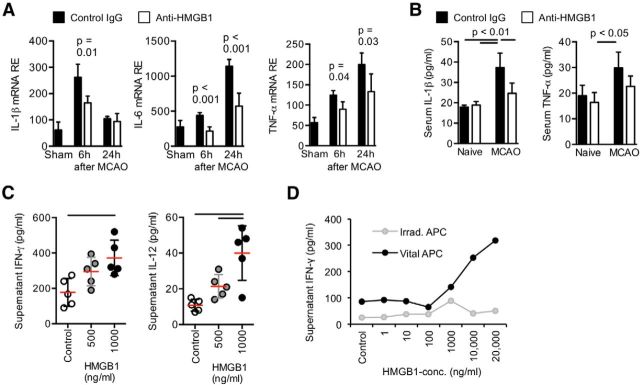
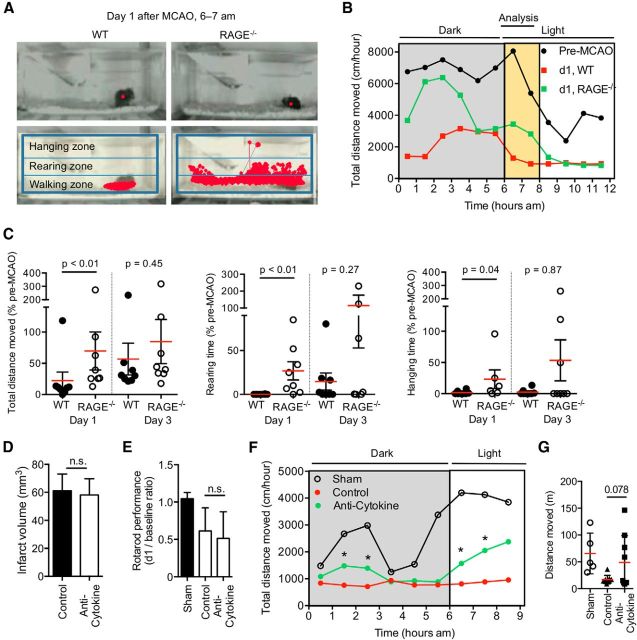
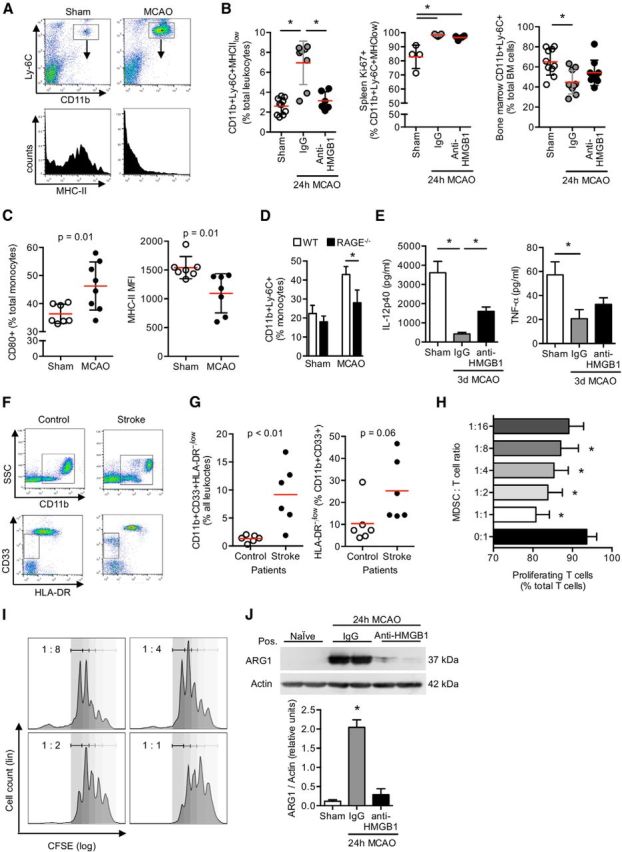
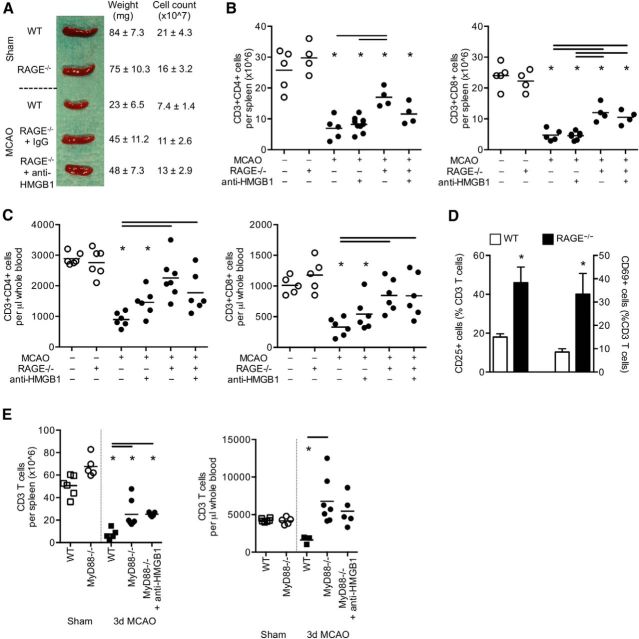
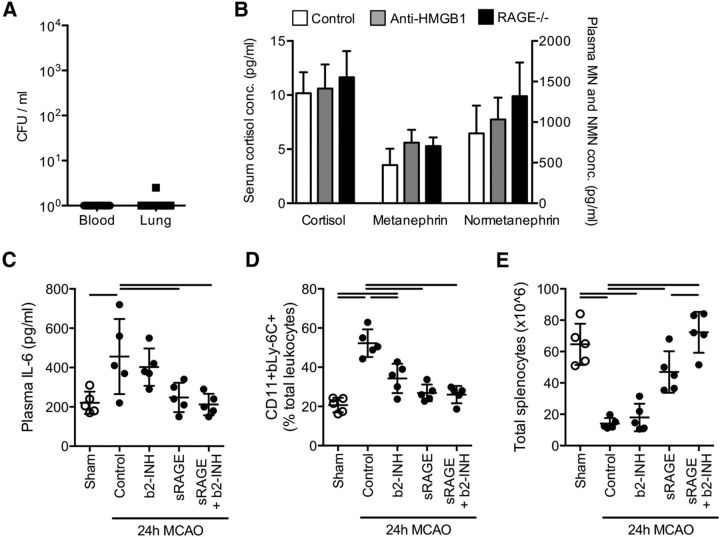
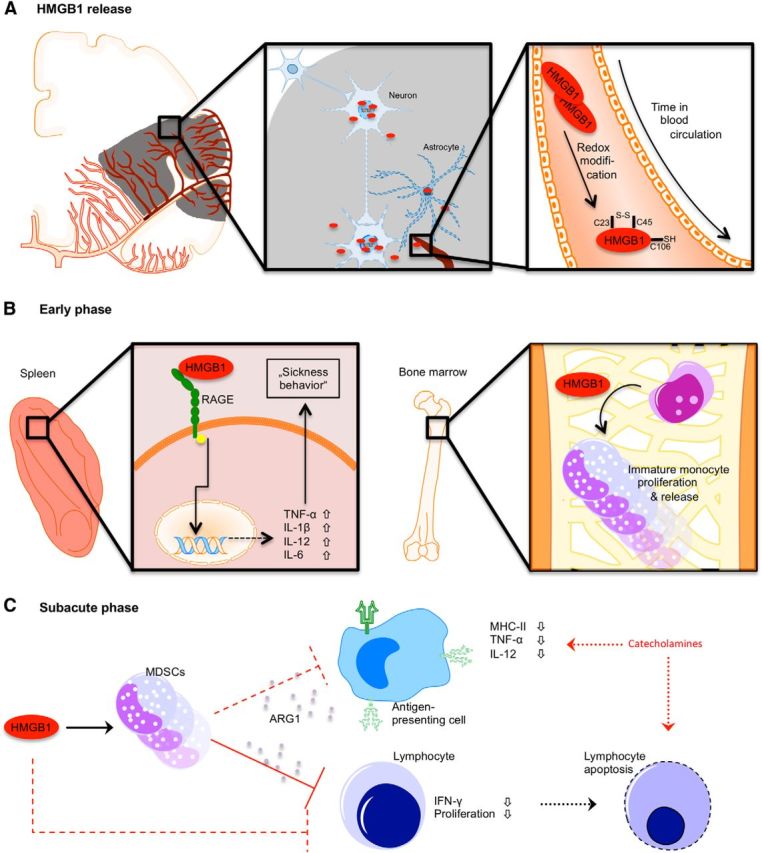
Acute brain lesions induce profound alterations of the peripheral immune response comprising the opposing phenomena of early immune activation and subsequent immunosuppression. The mechanisms underlying this brain-immune signaling are largely unknown. We used animal models for experimental brain ischemia as a paradigm of acute brain lesions and additionally investigated a large cohort of stroke patients. We analyzed release of HMGB1 isoforms by mass spectrometry and investigated its inflammatory potency and signaling pathways by immunological in vivo and in vitro techniques. Features of the complex behavioral sickness behavior syndrome were characterized by homecage behavior analysis. HMGB1 downstream signaling, particularly with RAGE, was studied in various transgenic animal models and by pharmacological blockade. Our results indicate that the cytokine-inducing, fully reduced isoform of HMGB1 was released from the ischemic brain in the hyperacute phase of stroke in mice and patients. Cytokines secreted in the periphery in response to brain injury induced sickness behavior, which could be abrogated by inhibition of the HMGB1-RAGE pathway or direct cytokine neutralization. Subsequently, HMGB1-release induced bone marrow egress and splenic proliferation of bone marrow-derived suppressor cells, inhibiting the adaptive immune responses in vivo and vitro. Furthermore, HMGB1-RAGE signaling resulted in functional exhaustion of mature monocytes and lymphopenia, the hallmarks of immune suppression after extensive ischemia. This study introduces the HMGB1-RAGE-mediated pathway as a key mechanism explaining the complex postischemic brain-immune interactions.
Keywords: HMGB1; RAGE; alarmins; immunomodulation; myeloid-derived suppressor cell; stroke.
Copyright © 2015 the authors 0270-6474/15/350583-16$15.00/0.
Figures
References
-
- Andrassy M, Volz HC, Igwe JC, Funke B, Eichberger SN, Kaya Z, Buss S, Autschbach F, Pleger ST, Lukic IK, Bea F, Hardt SE, Humpert PM, Bianchi ME, Mairbäurl H, Nawroth PP, Remppis A, Katus HA, Bierhaus A. High-mobility group box-1 in ischemia-reperfusion injury of the heart. Circulation. 2008;117:3216–3226. doi: 10.1161/CIRCULATIONAHA.108.769331. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical