

The origin and early evolution of metatherian mammals: the Cretaceous record
- PMID: 25589872
- PMCID: PMC4284630
- DOI: 10.3897/zookeys.465.8178
The origin and early evolution of metatherian mammals: the Cretaceous record
Abstract
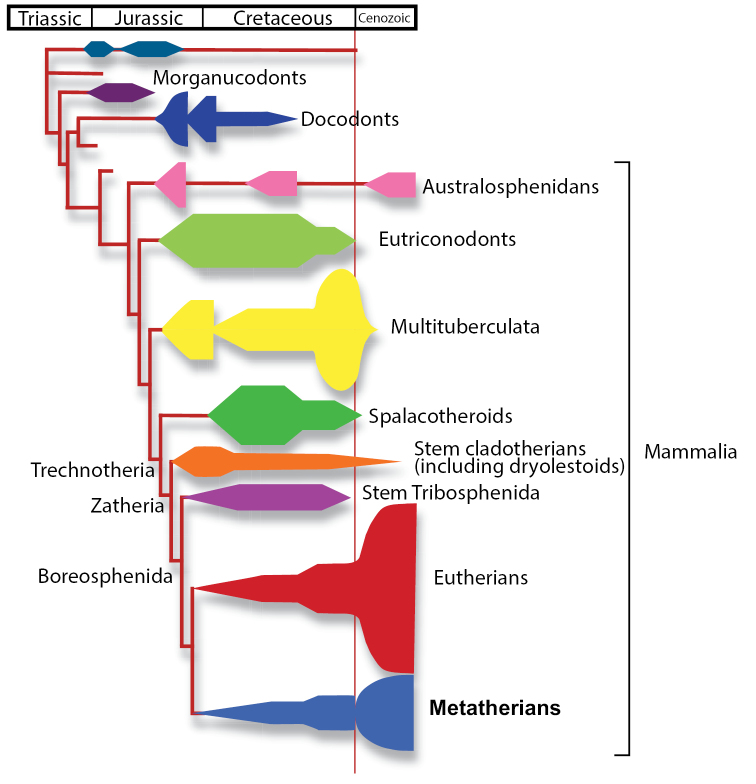
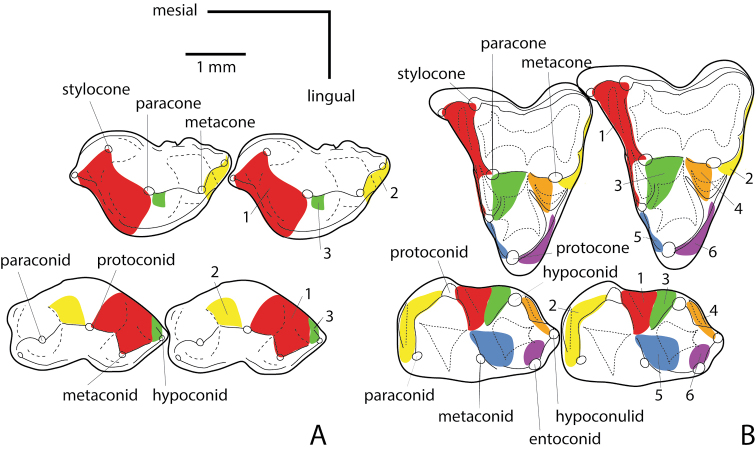
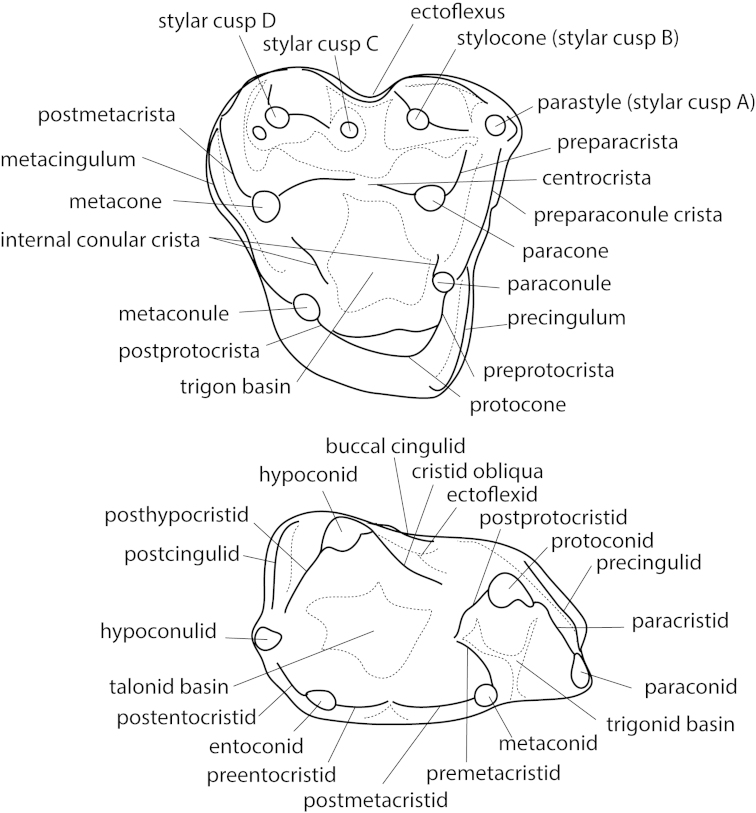
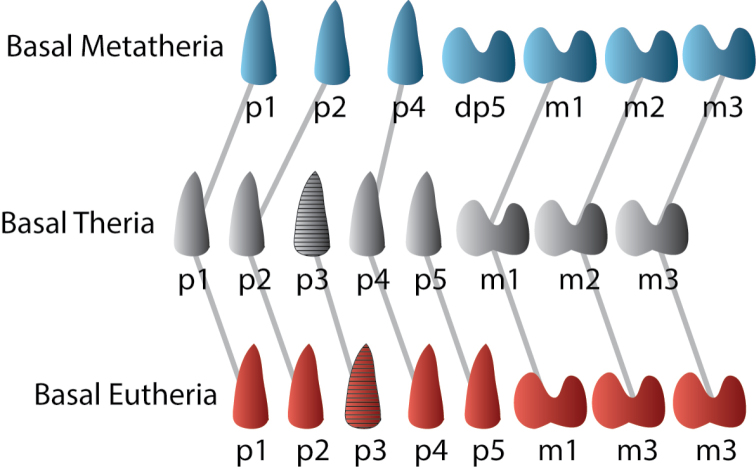
Metatherians, which comprise marsupials and their closest fossil relatives, were one of the most dominant clades of mammals during the Cretaceous and are the most diverse clade of living mammals after Placentalia. Our understanding of this group has increased greatly over the past 20 years, with the discovery of new specimens and the application of new analytical tools. Here we provide a review of the phylogenetic relationships of metatherians with respect to other mammals, discuss the taxonomic definition and diagnosis of Metatheria, outline the Cretaceous history of major metatherian clades, describe the paleobiology, biogeography, and macroevolution of Cretaceous metatherians, and provide a physical and climatic background of Cretaceous metatherian faunas. Metatherians are a clade of boreosphendian mammals that must have originated by the Late Jurassic, but the first unequivocal metatherian fossil is from the Early Cretaceous of Asia. Metatherians have the distinctive tightly interlocking occlusal molar pattern of tribosphenic mammals, but differ from Eutheria in their dental formula and tooth replacement pattern, which may be related to the metatherian reproductive process which includes an extended period of lactation followed by birth of extremely altricial young. Metatherians were widespread over Laurasia during the Cretaceous, with members present in Asia, Europe, and North America by the early Late Cretaceous. In particular, they were taxonomically and morphologically diverse and relatively abundant in the Late Cretaceous of western North America, where they have been used to examine patterns of biogeography, macroevolution, diversification, and extinction through the Late Cretaceous and across the Cretaceous-Paleogene (K-Pg) boundary. Metatherian diversification patterns suggest that they were not strongly affected by a Cretaceous Terrestrial Revolution, but they clearly underwent a severe extinction across the K-Pg boundary.
Keywords: Boreosphenida; Cretaceous; Deltatheroida; Mammalia; Marsupialiformes; Metatheria; biogeography; dentition; macroevolution; osteology; paleobiology; paleoenvironment; phylogeny.
Figures
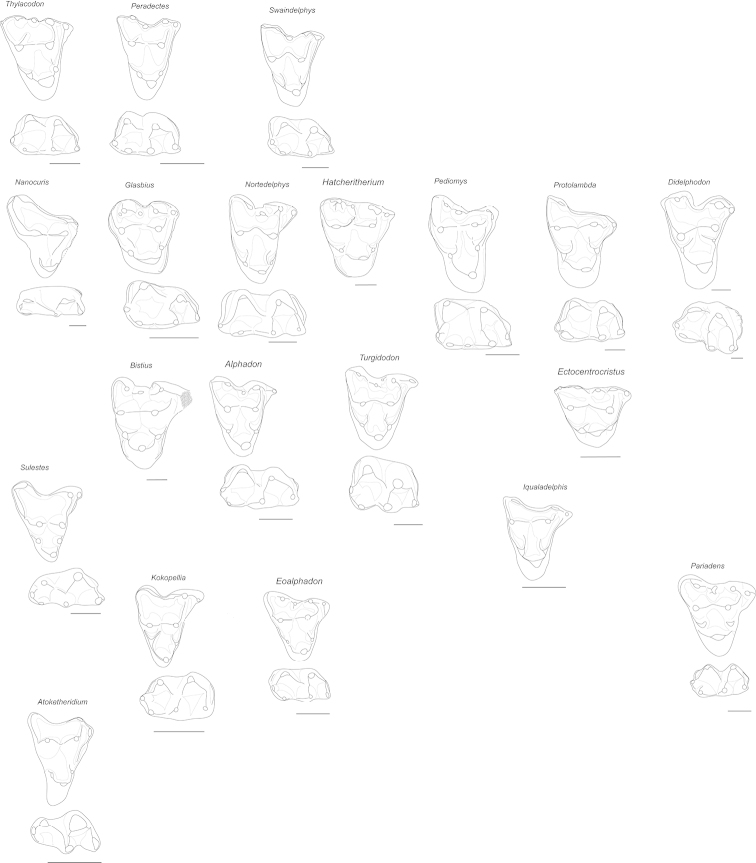
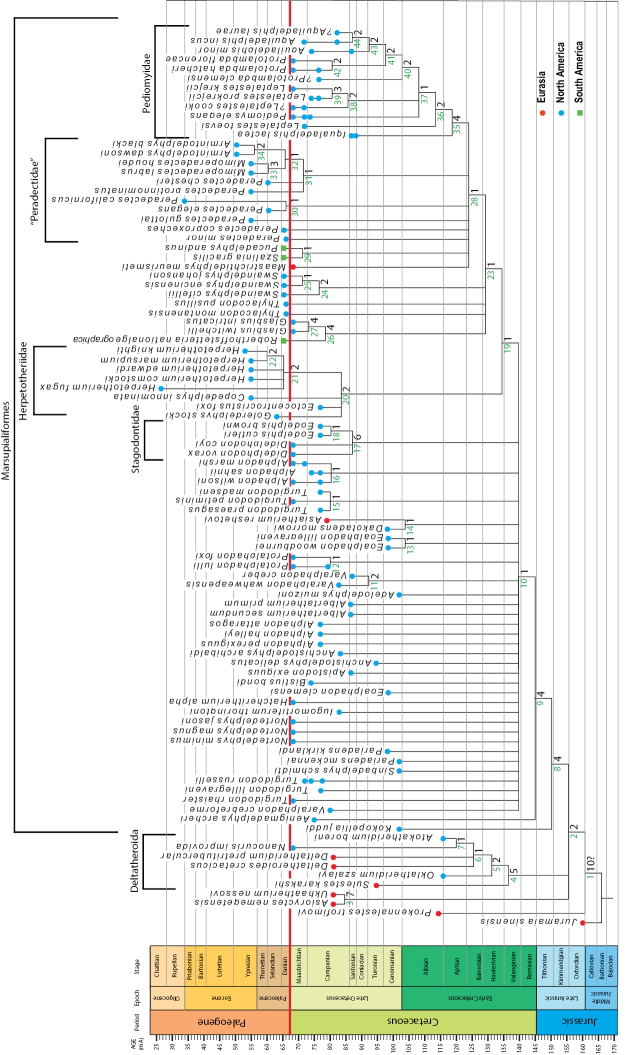
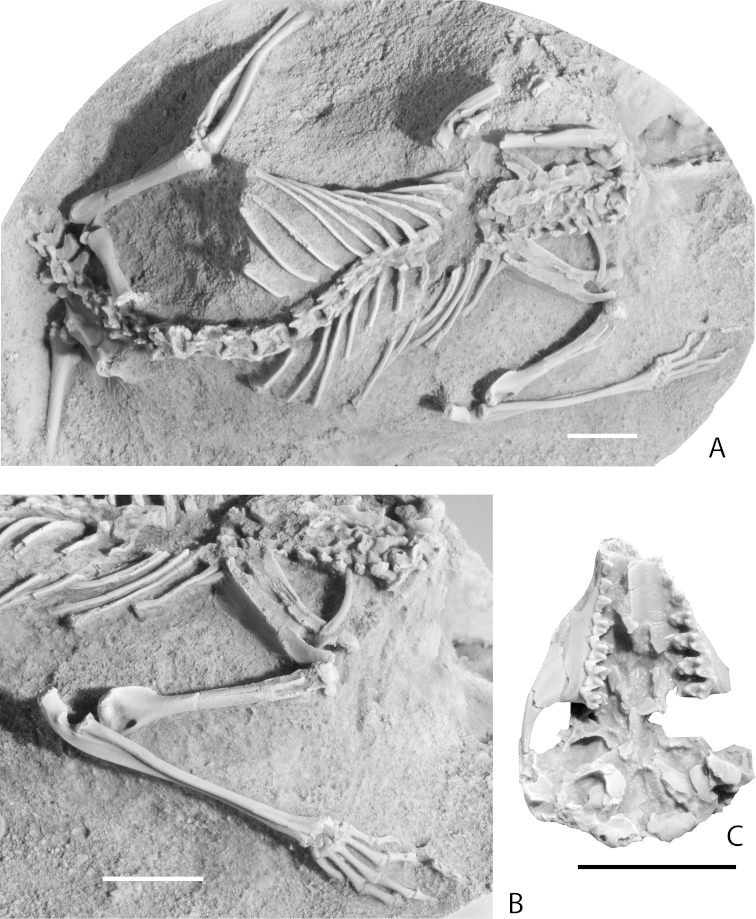
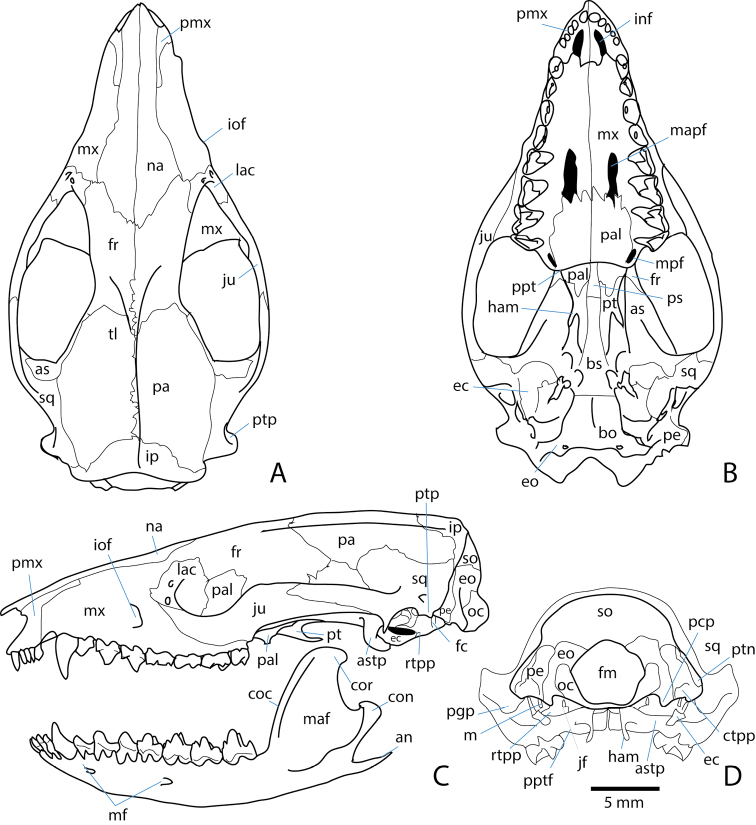
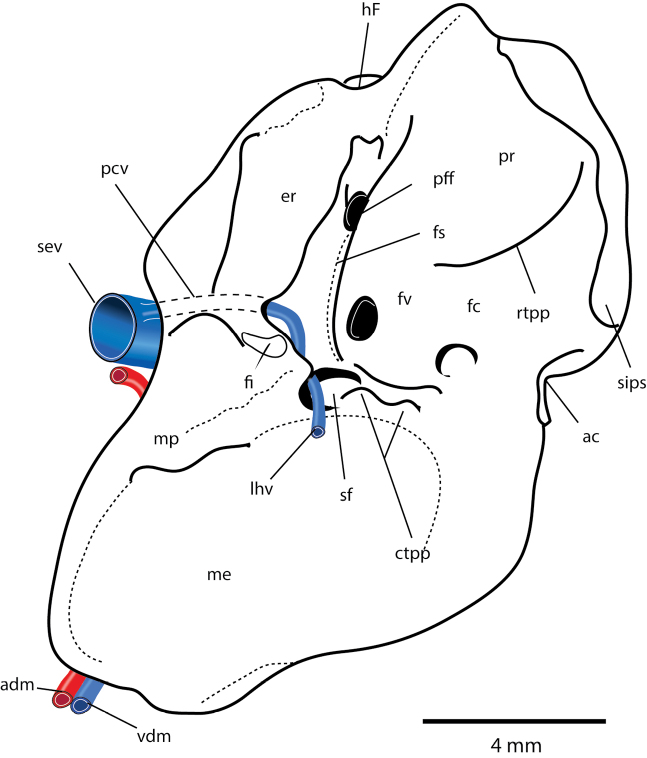
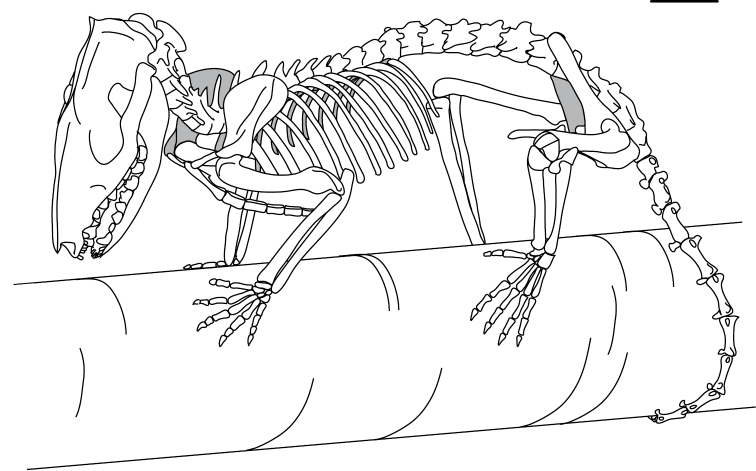
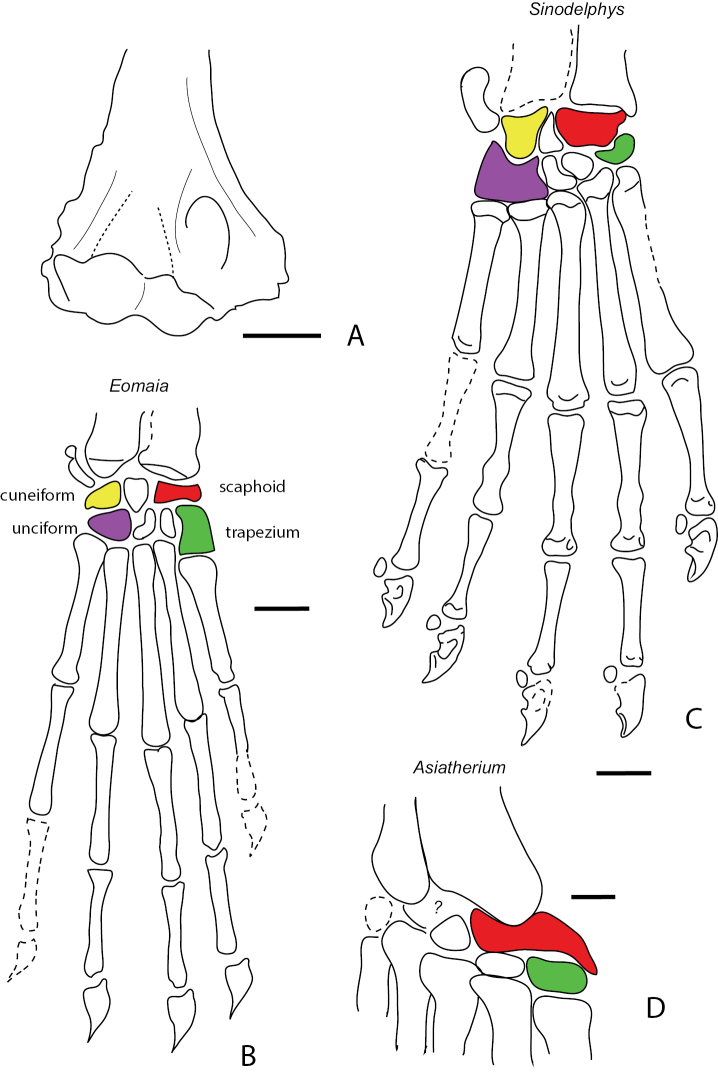
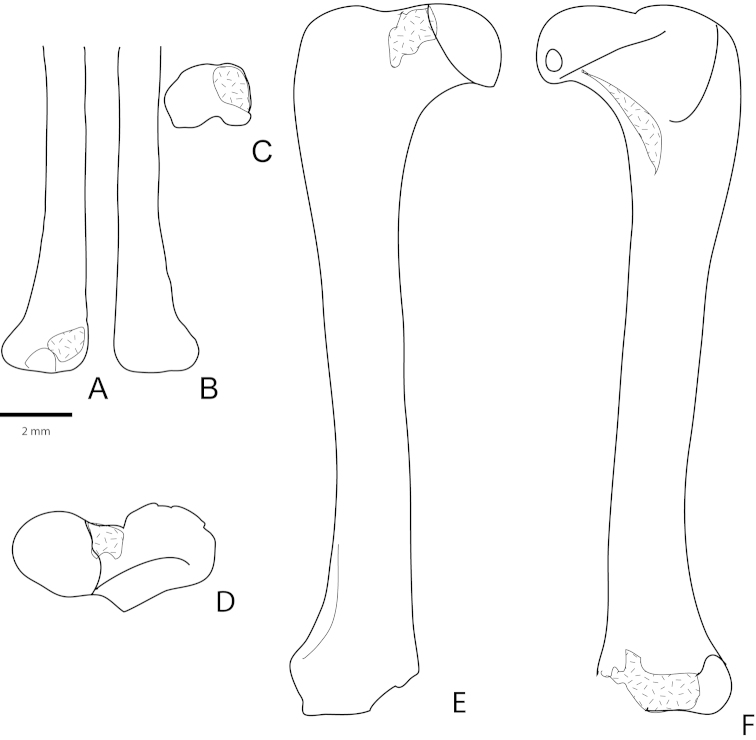
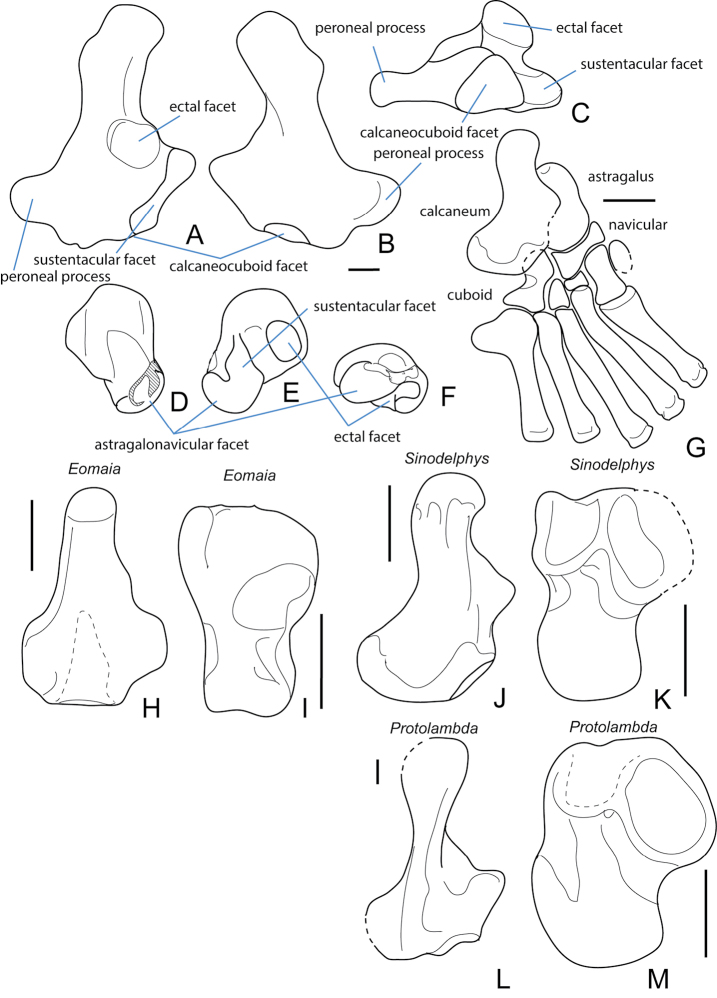
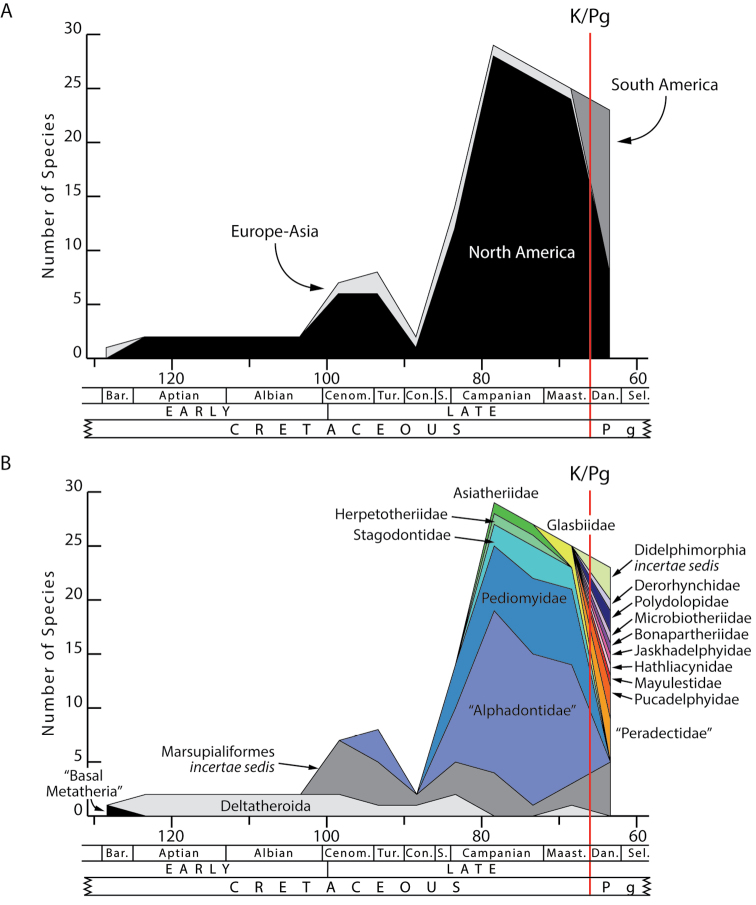
References
-
- Archibald JD. (1982) A study of Mammalia and geology across the Cretaceous-Tertiary boundary in Garfield County, Montana. University of California Publications in Geological Sciences 122: 1–286.
-
- Archibald JD. (1987) Late Cretaceous (Judithian and Edmontonian) vertebrates and geology of the Williams Fork Formation, N. W. Colorado. Fourth Symposium on Mesozoic Terrestrial Ecosystems; Occasional Paper of the Tyrrell Museum of Palaeontology 3: 7–11.
-
- Archibald JD, Averianov AO. (2005) Mammalian faunal succession in the Cretaceous of the Kyzylkum Desert. Journal of Mammalian Evolution 12: 9–22. doi: 10.1007/s10914-005-4867-3 - DOI
-
- Archibald JD, Zhang Y, Harper T, Cifelli RL. (2011) Protungulatum, confirmed Cretaceous occurrence of an otherwise Paleocene eutherian (Placental?) mammal. Journal of Mammalian Evolution 18: 153–161. doi: 10.1007/s10914-011-9162-1 - DOI
-
- Argot C. (2001) Functional-adaptive anatomy of the forelimb in the Didelphidae and the paleobiology of the Paleocene marsupials Mayulestes ferox and Pucadelphys andinus. Journal of Morphology 247: 51–79. doi: 10.1002/1097-4687(200101)247:1<51::AID-JMOR1003>3.0.CO;2-# - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous