

Glial lipid droplets and ROS induced by mitochondrial defects promote neurodegeneration
- PMID: 25594180
- PMCID: PMC4377295
- DOI: 10.1016/j.cell.2014.12.019
Glial lipid droplets and ROS induced by mitochondrial defects promote neurodegeneration
Abstract
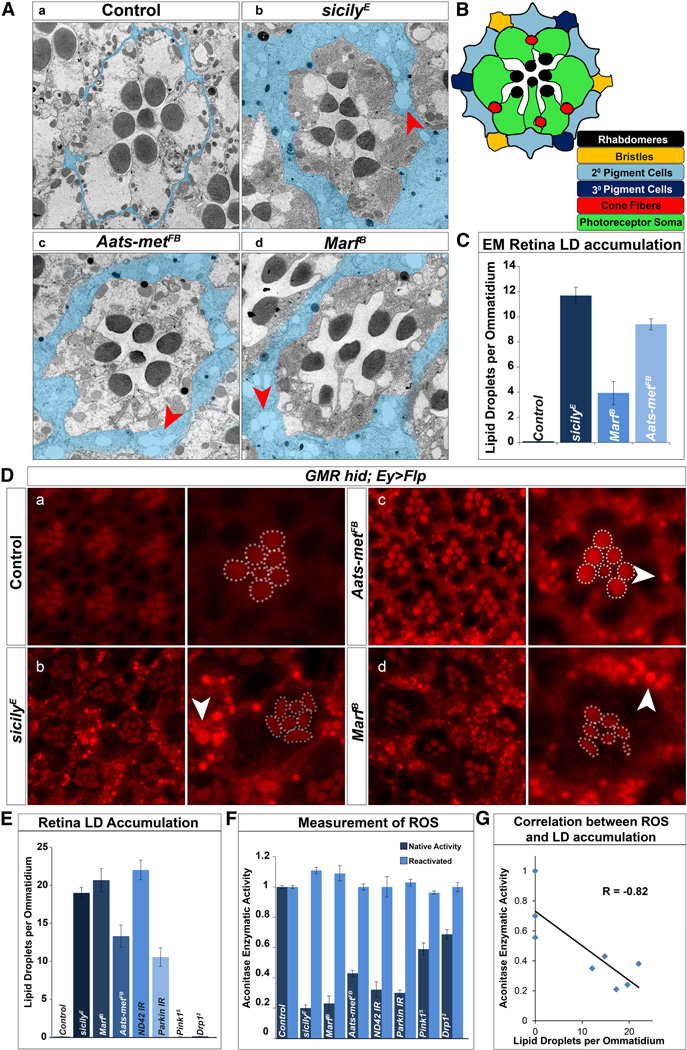
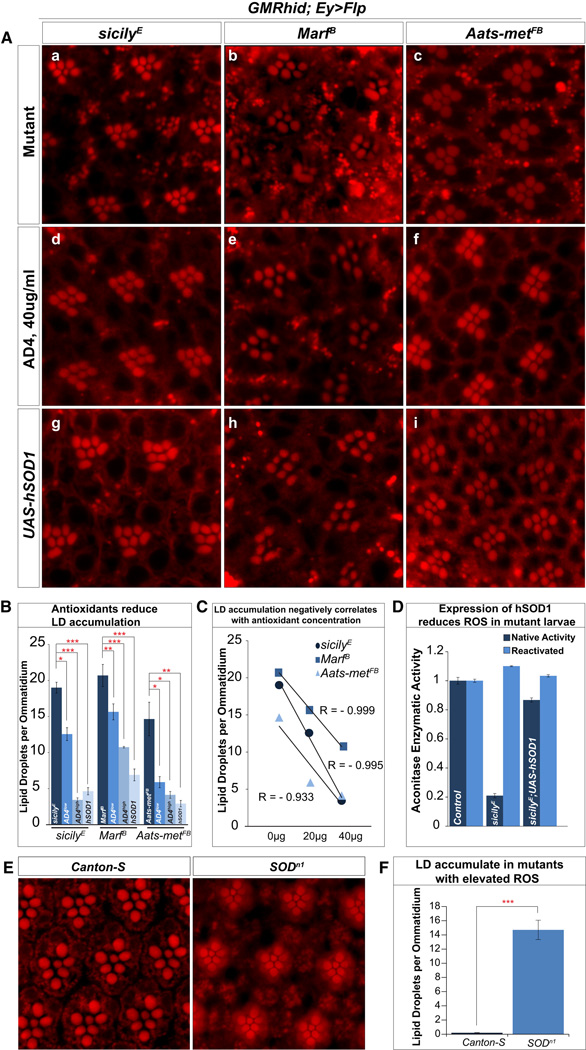
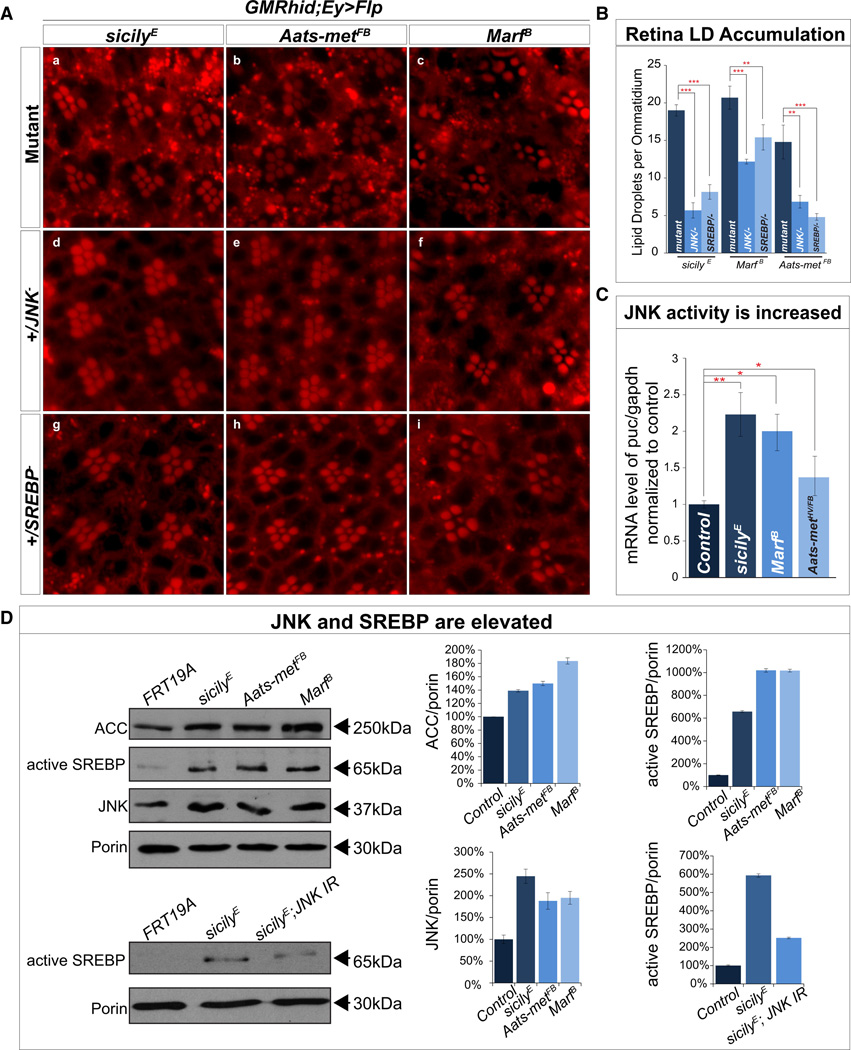
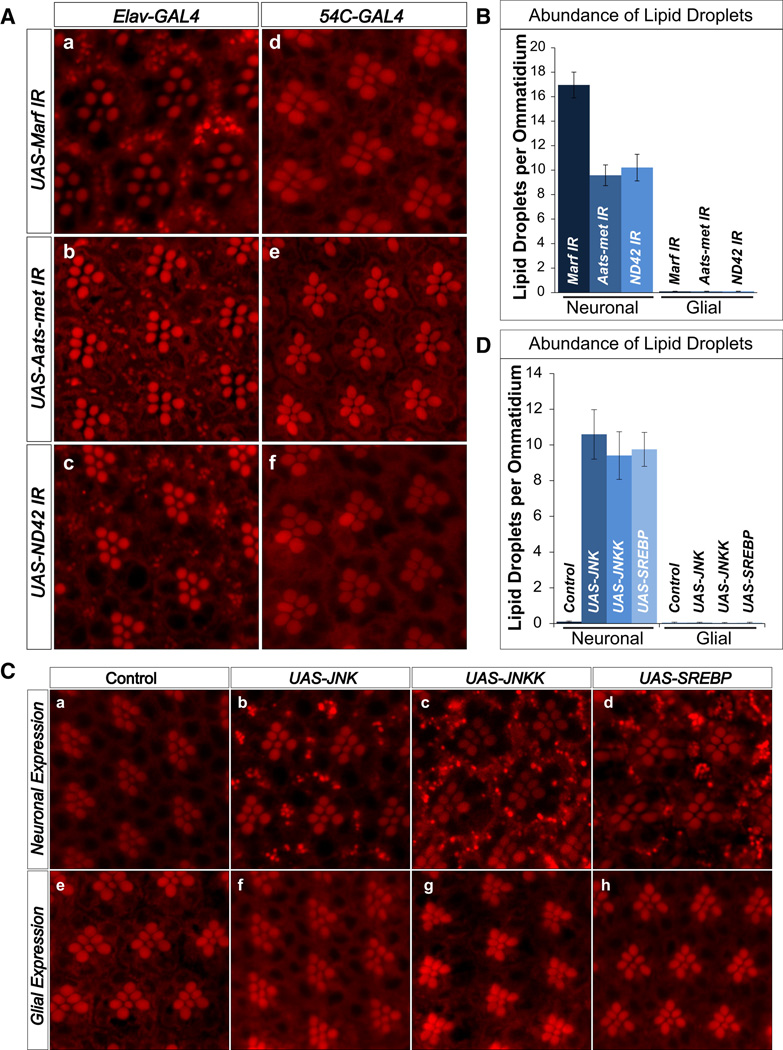
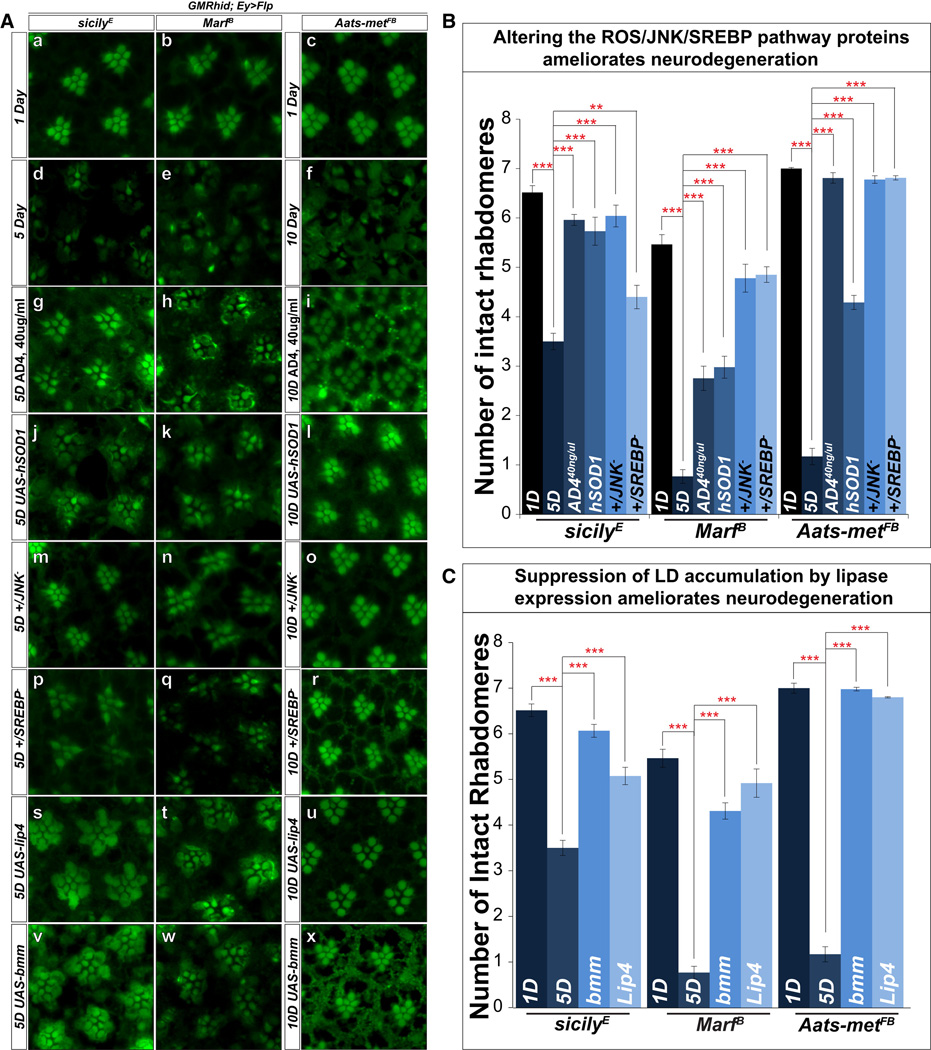
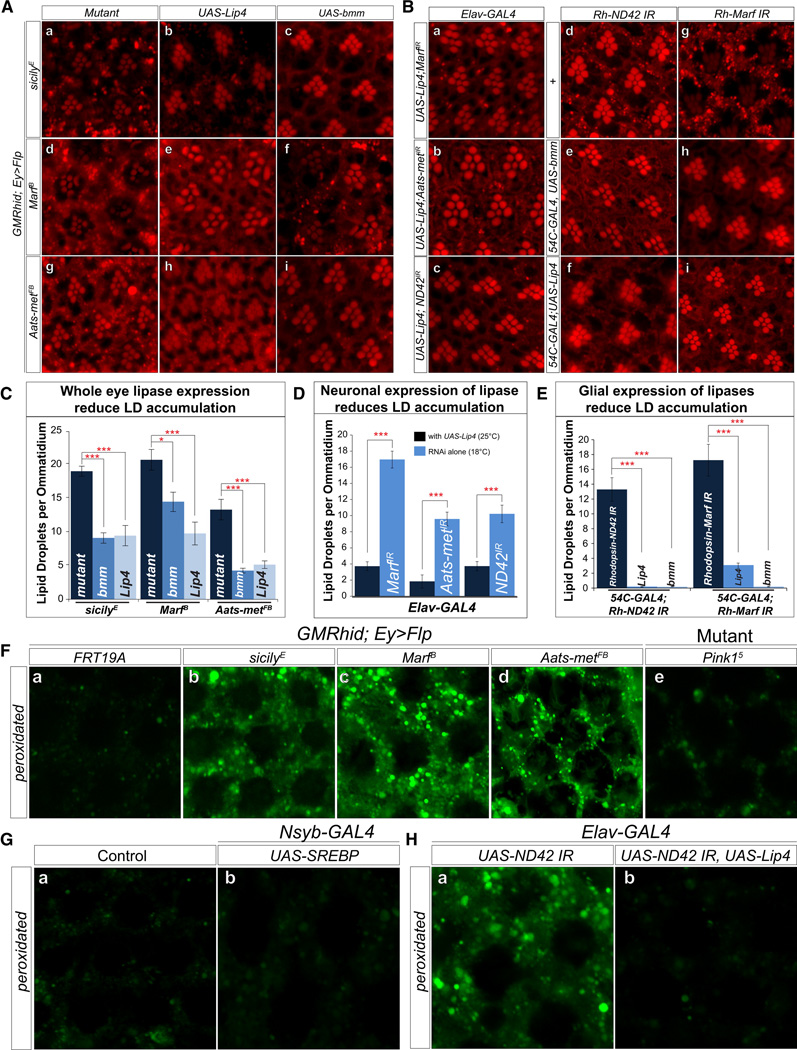
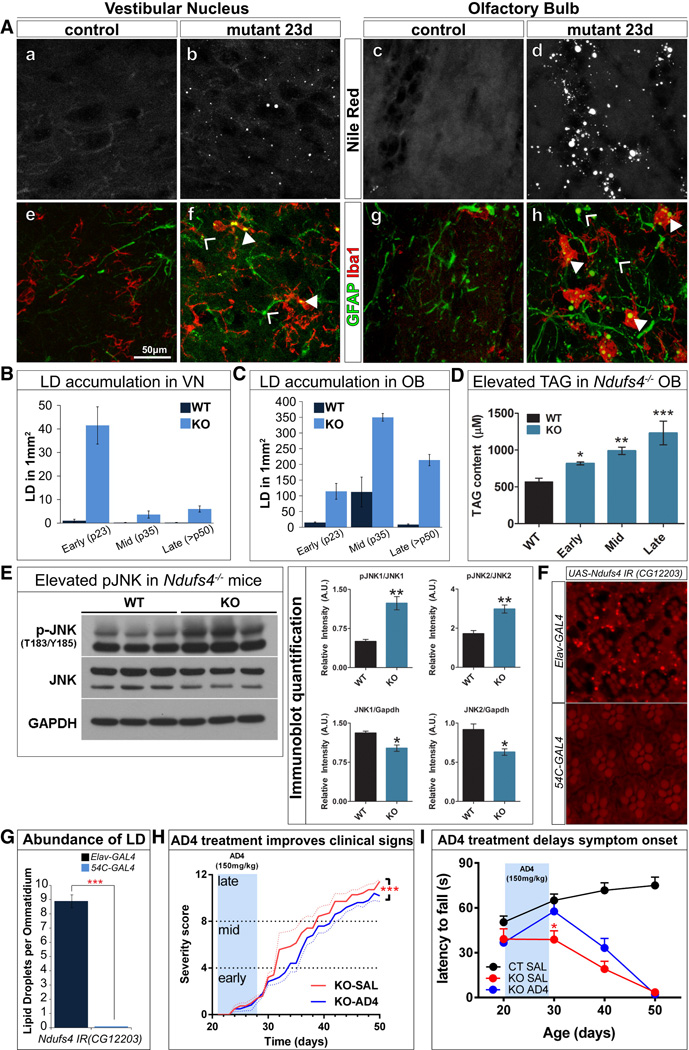
Reactive oxygen species (ROS) and mitochondrial defects in neurons are implicated in neurodegenerative disease. Here, we find that a key consequence of ROS and neuronal mitochondrial dysfunction is the accumulation of lipid droplets (LD) in glia. In Drosophila, ROS triggers c-Jun-N-terminal Kinase (JNK) and Sterol Regulatory Element Binding Protein (SREBP) activity in neurons leading to LD accumulation in glia prior to or at the onset of neurodegeneration. The accumulated lipids are peroxidated in the presence of ROS. Reducing LD accumulation in glia and lipid peroxidation via targeted lipase overexpression and/or lowering ROS significantly delays the onset of neurodegeneration. Furthermore, a similar pathway leads to glial LD accumulation in Ndufs4 mutant mice with neuronal mitochondrial defects, suggesting that LD accumulation following mitochondrial dysfunction is an evolutionarily conserved phenomenon, and represents an early, transient indicator and promoter of neurodegenerative disease.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
References
-
- Amer J, Atlas D, Fibach E. N-acetylcysteine amide (AD4) attenuates oxidative stress in betathalassemia blood cells. Biochim. Biophys. Acta. 2008;1780:249–255. - PubMed
-
- Assouline Z, Jambou M, Rio M, Bole-Feysot C, de LP, Barnerias C, Desguerre I, Bonnemains C, Guillermet C, Steffann J, Munnich A, Bonnefont JP, Rotig A, Lebre AS. A constant and similar assembly defect of mitochondrial respiratory chain complex I allows rapid identification of NDUFS4 mutations in patients with Leigh syndrome. Biochim. Biophys. Acta. 2012;1822:1062–1069. - PubMed
-
- Barnham KJ, Masters CL, Bush AI. Neurodegenerative diseases and oxidative stress. Nat. Rev. Drug Discov. 2004;3:205–214. - PubMed
-
- Bayat V, Thiffault I, Jaiswal M, Tetreault M, Donti T, Sasarman F, Bernard G, Demers-Lamarche J, Dicaire MJ, Mathieu J, Vanasse M, Bouchard JP, Rioux MF, Lourenco CM, Li Z, Haueter C, Shoubridge EA, Graham BH, Brais B, Bellen HJ. Mutations in the mitochondrial methionyl-tRNA synthetase cause a neurodegenerative phenotype in flies and a recessive ataxia (ARSAL) in humans. PLoS. Biol. 2012;10:e1001288. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
