

Reduction of the temperature sensitivity of Halomonas hydrothermalis by iron starvation combined with microaerobic conditions
- PMID: 25595757
- PMCID: PMC4345356
- DOI: 10.1128/AEM.03639-14
Reduction of the temperature sensitivity of Halomonas hydrothermalis by iron starvation combined with microaerobic conditions
Abstract
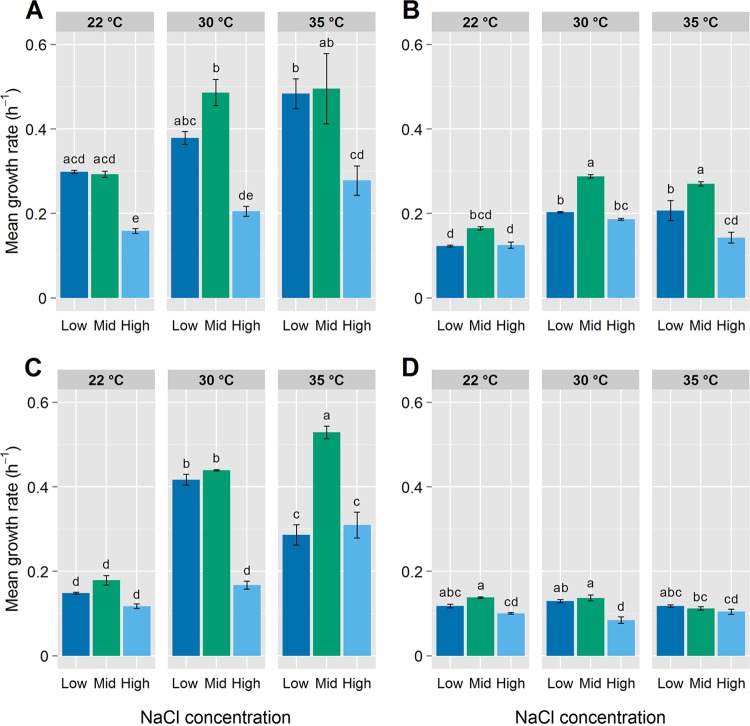
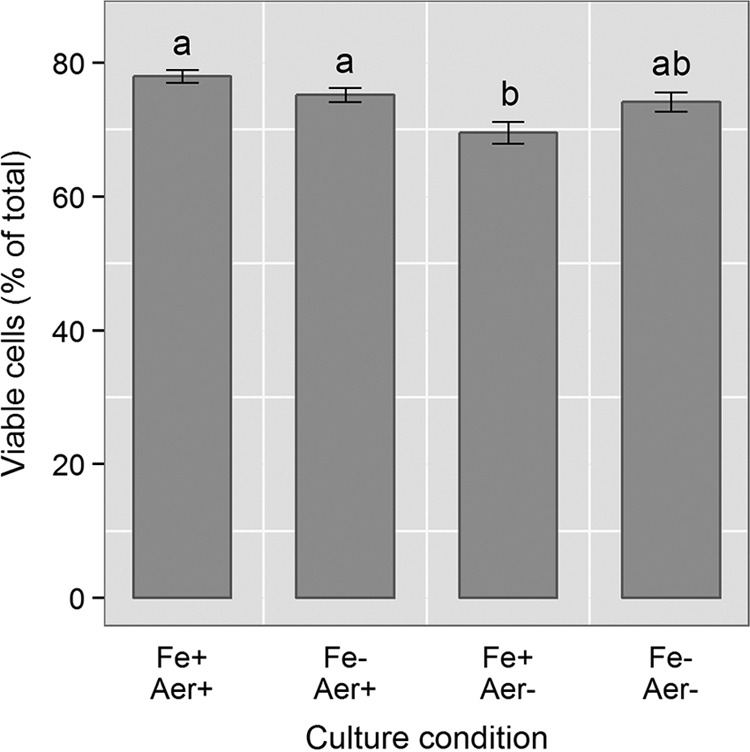
The limits to biological processes on Earth are determined by physicochemical parameters, such as extremes of temperature and low water availability. Research into microbial extremophiles has enhanced our understanding of the biophysical boundaries which define the biosphere. However, there remains a paucity of information on the degree to which rates of microbial multiplication within extreme environments are determined by the availability of specific chemical elements. Here, we show that iron availability and the composition of the gaseous phase (aerobic versus microaerobic) determine the susceptibility of a marine bacterium, Halomonas hydrothermalis, to suboptimal and elevated temperature and salinity by impacting rates of cell division (but not viability). In particular, iron starvation combined with microaerobic conditions (5% [vol/vol] O2, 10% [vol/vol] CO2, reduced pH) reduced sensitivity to temperature across the 13°C range tested. These data demonstrate that nutrient limitation interacts with physicochemical parameters to determine biological permissiveness for extreme environments. The interplay between resource availability and stress tolerance, therefore, may shape the distribution and ecology of microorganisms within Earth's biosphere.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
Similar articles
-
Astrobiology as a framework for investigating antibiotic susceptibility: a study of Halomonas hydrothermalis.J R Soc Interface. 2017 Jan;14(126):20160942. doi: 10.1098/rsif.2016.0942. J R Soc Interface. 2017. PMID: 28123098 Free PMC article.
-
Synchronous effects of temperature, hydrostatic pressure, and salinity on growth, phospholipid profiles, and protein patterns of four Halomonas species isolated from deep-sea hydrothermal-vent and sea surface environments.Appl Environ Microbiol. 2004 Oct;70(10):6220-9. doi: 10.1128/AEM.70.10.6220-6229.2004. Appl Environ Microbiol. 2004. PMID: 15466569 Free PMC article.
-
Efficient secretion of (R)-3-hydroxybutyric acid from Halomonas sp. KM-1 by nitrate fed-batch cultivation with glucose under microaerobic conditions.Bioresour Technol. 2014 Mar;156:400-3. doi: 10.1016/j.biortech.2014.01.073. Epub 2014 Jan 27. Bioresour Technol. 2014. PMID: 24503050
-
The limits for life under multiple extremes.Trends Microbiol. 2013 Apr;21(4):204-12. doi: 10.1016/j.tim.2013.01.006. Epub 2013 Feb 27. Trends Microbiol. 2013. PMID: 23453124 Review.
-
Limitations of microbial iron reduction under extreme conditions.FEMS Microbiol Rev. 2022 Nov 2;46(6):fuac033. doi: 10.1093/femsre/fuac033. FEMS Microbiol Rev. 2022. PMID: 35849069 Free PMC article. Review.
Cited by
-
The UK Centre for Astrobiology: A Virtual Astrobiology Centre. Accomplishments and Lessons Learned, 2011-2016.Astrobiology. 2018 Feb;18(2):224-243. doi: 10.1089/ast.2017.1713. Epub 2018 Jan 29. Astrobiology. 2018. PMID: 29377716 Free PMC article.
-
Is there a common water-activity limit for the three domains of life?ISME J. 2015 Jun;9(6):1333-51. doi: 10.1038/ismej.2014.219. Epub 2014 Dec 12. ISME J. 2015. PMID: 25500507 Free PMC article.
-
Stress tolerance and virulence of insect-pathogenic fungi are determined by environmental conditions during conidial formation.Curr Genet. 2015 Aug;61(3):383-404. doi: 10.1007/s00294-015-0477-y. Epub 2015 Mar 20. Curr Genet. 2015. PMID: 25791499
-
Where the microbes aren't.FEMS Microbiol Rev. 2025 Jan 14;49:fuae034. doi: 10.1093/femsre/fuae034. FEMS Microbiol Rev. 2025. PMID: 39725411 Free PMC article. Review.
-
Aerobically respiring prokaryotic strains exhibit a broader temperature-pH-salinity space for cell division than anaerobically respiring and fermentative strains.J R Soc Interface. 2015 Sep 6;12(110):0658. doi: 10.1098/rsif.2015.0658. J R Soc Interface. 2015. PMID: 26354829 Free PMC article.
References
-
- Lammer H, Bredehöft JH, Coustenis A, Khodachenko ML, Kaltenegger L, Grasset O, Prieur D, Raulin F, Ehrenfreund P, Yamauchi M, Wahlund J-E, Grießmeier J-M, Stangl G, Cockell CS, Kulikov YN, Grenfell JL, Rauer H. 2009. What makes a planet habitable? Astron Astrophys Rev 17:181–249. doi:10.1007/s00159-009-0019-z. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
