

Natural IgM prevents autoimmunity by enforcing B cell central tolerance induction
- PMID: 25595791
- PMCID: PMC4324358
- DOI: 10.4049/jimmunol.1401880
Natural IgM prevents autoimmunity by enforcing B cell central tolerance induction
Abstract
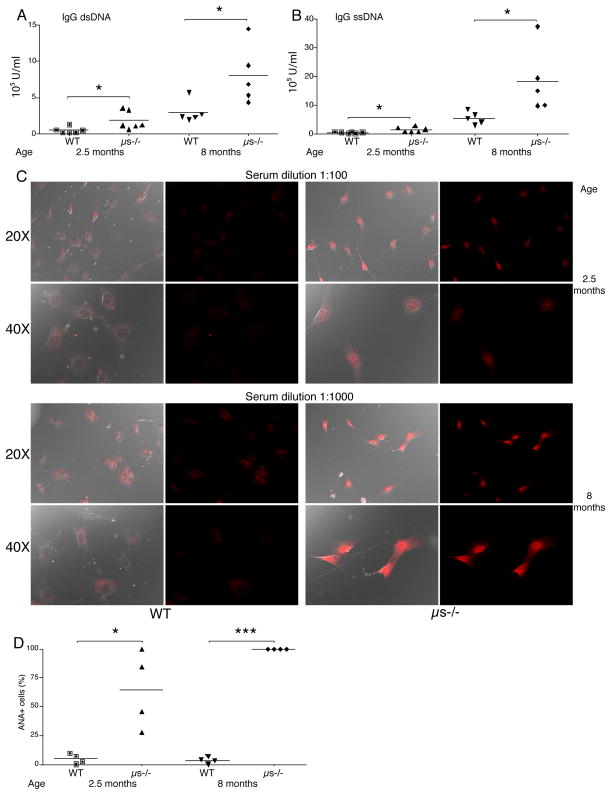
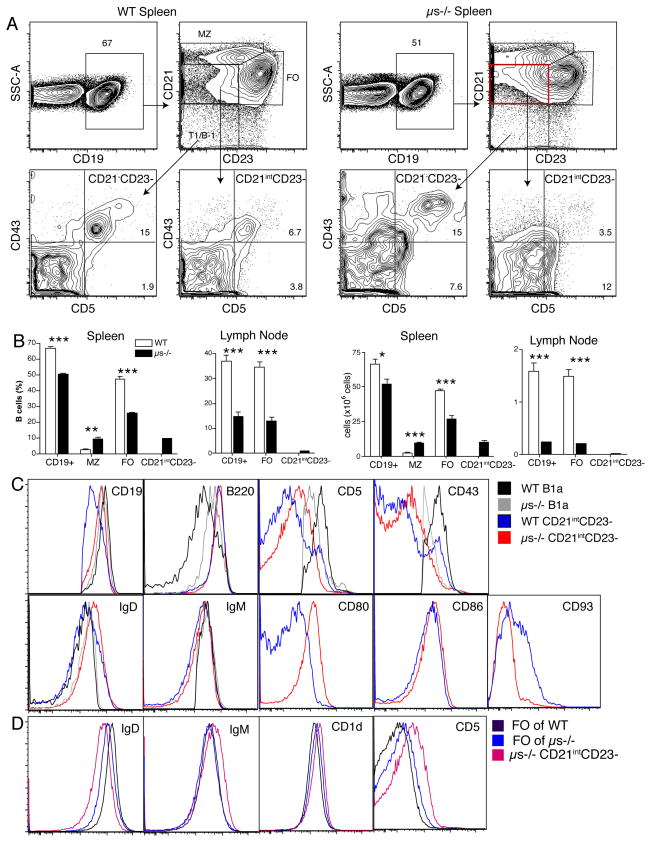
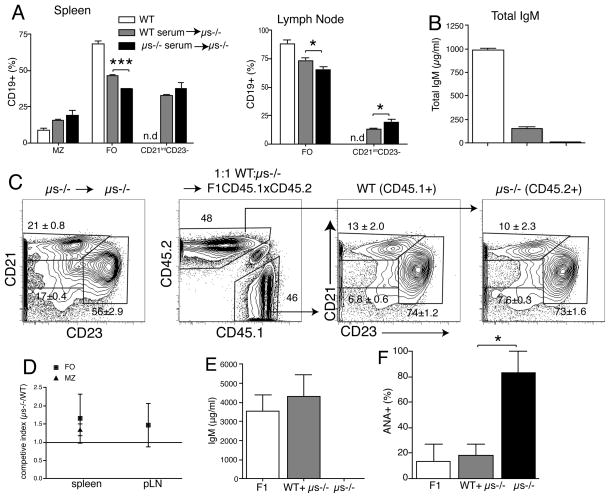
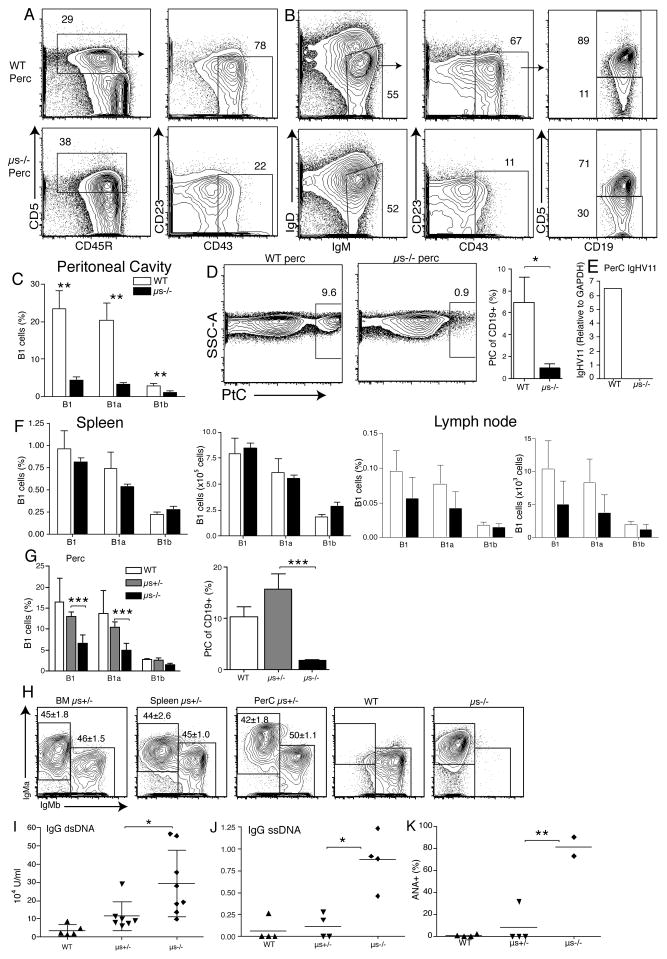
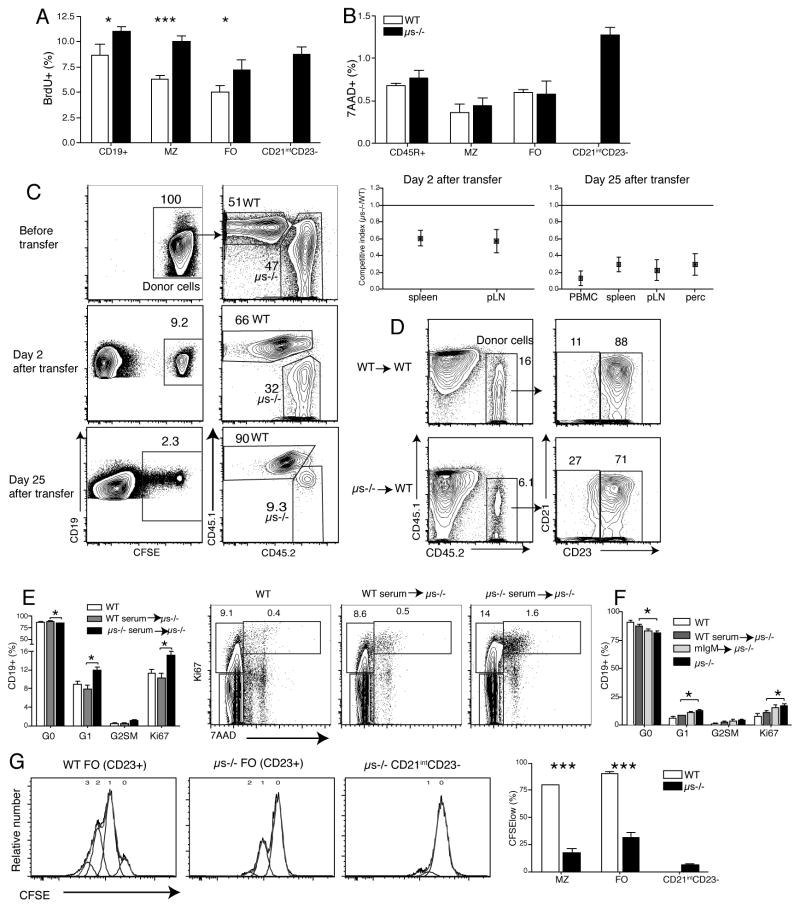
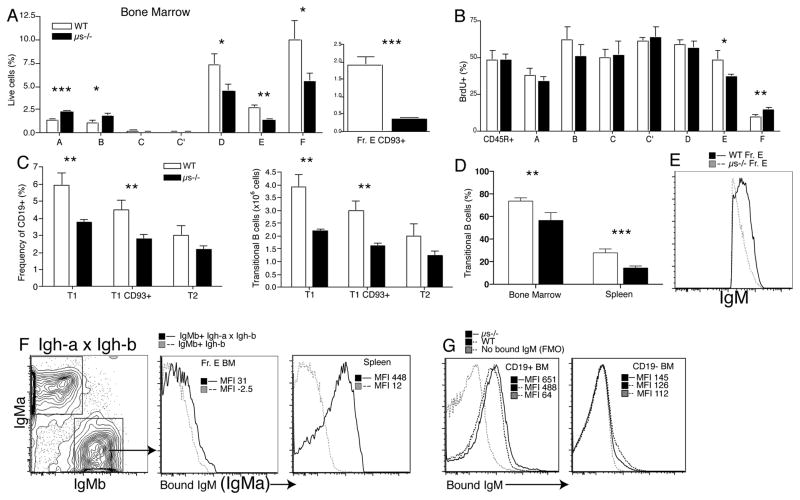
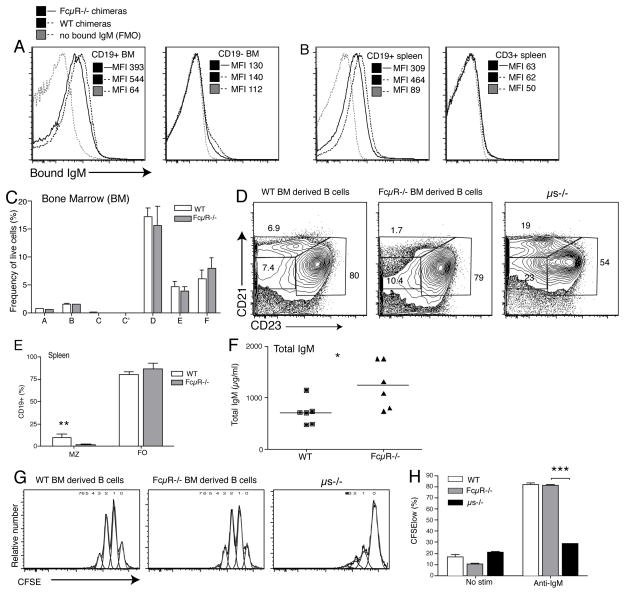
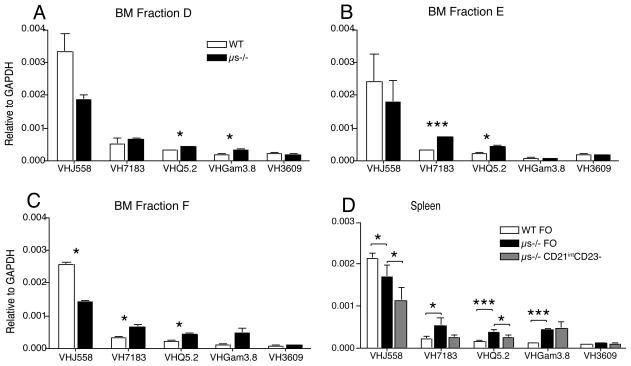
It is unclear why selective deficiency in secreted (s)IgM causes Ab-mediated autoimmunity. We demonstrate that sIgM is required for normal B cell development and selection. The CD5(+) B cells that were previously shown to accumulate in body cavities of sIgM(-/-) mice are not B-1a cells, but CD19(int), CD43(-), short-lived, BCR signaling-unresponsive anergic B-2 cells. Body cavity B-1 cells were >10-fold reduced, including VH11(+) and phosphotidylcholine-specific B-1a cells, whereas splenic B-1 cells were unaffected and marginal zone B cells increased. Follicular B cells had higher turnover rates, survived poorly after adoptive transfer, and were unresponsiveness to BCR stimulation in vitro. sIgM bound to B cell precursors and provided a positive signal to overcome a block at the pro/pre-B stage and during IgVH repertoire selection. Polyclonal IgM rescued B cell development and returned autoantibody levels to near normal. Thus, natural IgM deficiency causes primary autoimmune disease by altering B cell development, selection, and central tolerance induction.
Copyright © 2015 by The American Association of Immunologists, Inc.
Figures
References
-
- Ehrenstein MR, Notley CA. The importance of natural IgM: scavenger, protector and regulator. Nature reviews Immunology. 2010;10:778–786. - PubMed
-
- Hooijkaas H, van der Linde-Preesman AA, Benne S, Benner R. Frequency analysis of the antibody specificity repertoire of mitogen-reactive B cells and “spontaneously” occurring “background” plaque-forming cells in nude mice. Cellular immunology. 1985;92:154–162. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
