

Amyloid accumulation is a late event in sporadic Alzheimer's disease-like pathology in nontransgenic rats
- PMID: 25595891
- PMCID: PMC4359302
- DOI: 10.18632/oncotarget.2751
Amyloid accumulation is a late event in sporadic Alzheimer's disease-like pathology in nontransgenic rats
Abstract
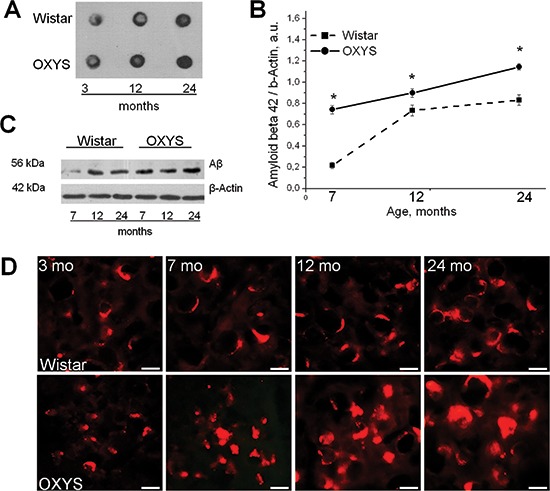
The amyloid cascade hypothesis posits that deposition of the amyloid β (Aβ) peptide in the brain is a key event in the initiation of Alzheimer's disease (AD). Nonetheless, it now seems increasingly unlikely that amyloid toxicity is the cause of sporadic AD, which leads to cognitive decline. Here, using accelerated-senescence nontransgenic OXYS rats, we confirmed that aggregation of Aβ is a later event in AD-like pathology. We showed that an age-dependent increase in the levels of Aβ₁₋₄₂ and extracellular Aβ deposits in the brain of OXYS rats occur later than do synaptic losses, neuronal cell death, mitochondrial structural abnormalities, and hyperphosphorylation of the tau protein. We identified the variants of the genes that are strongly associated with the risk of either late-onset or early-onset AD, including App, Apoe4, Bace1, Psen1, Psen2, and Picalm. We found that in OXYS rats nonsynonymous SNPs were located only in the genes Casp3 and Sorl1. Thus, we present proof that OXYS rats may be a model of sporadic AD. It is possible that multiple age-associated pathological processes may precede the toxic amyloid accumulation, which in turn triggers the final stage of the sporadic form of AD and becomes a hallmark event of the disease.
Conflict of interest statement
The authors declare that they have no competing financial interests.
Figures





References
-
- Morley JE, Armbrecht HJ, Farr SA, Kumar VB. The senescence accelerated mouse (SAMP8) as a model for oxidative stress and Alzheimer's disease. Biochim Biophys Acta. 2012;1822:650–656. - PubMed
-
- Drachman DA. The amyloid hypothesis, time to move on: Amyloid is the downstream result, not cause, of Alzheimer's disease. Alzheimers Dement. 2014;3:372–380. - PubMed
-
- Morley JE, Farr SA. The role of amyloid-beta in the regulation of memory. Biochem Pharmacol. 2014;88:479–485. - PubMed
-
- Krstic D, Knuesel I. Deciphering the mechanism underlying late-onset Alzheimer disease. Nat Rev Neurol. 2013;9:25–34. - PubMed
-
- Castellano JM, Deane R, Gottesdiener AJ, Verghese PB, Stewart FR, West T, Paoletti AC, Kasper TR, DeMattos RB, Zlokovic BV, Holtzman DM. Low-density lipoprotein receptor overexpression enhances the rate of brain-to-blood Aβ clearance in a mouse model of β-amyloidosis. Proc Natl Acad Sci U S A. 2012;109:15502–15507. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
