

Cocaine-seeking is associated with PKC-dependent reduction of excitatory signaling in accumbens shell D2 dopamine receptor-expressing neurons
- PMID: 25596492
- PMCID: PMC4346508
- DOI: 10.1016/j.neuropharm.2015.01.002
Cocaine-seeking is associated with PKC-dependent reduction of excitatory signaling in accumbens shell D2 dopamine receptor-expressing neurons
Abstract
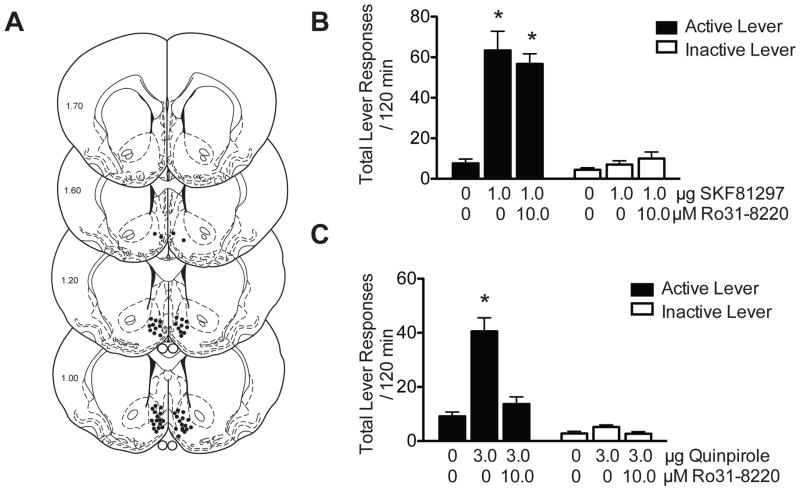
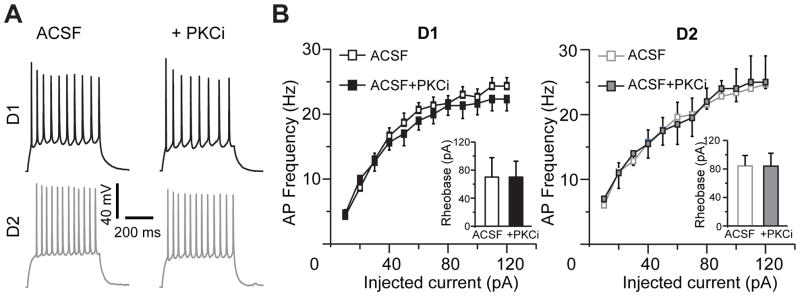
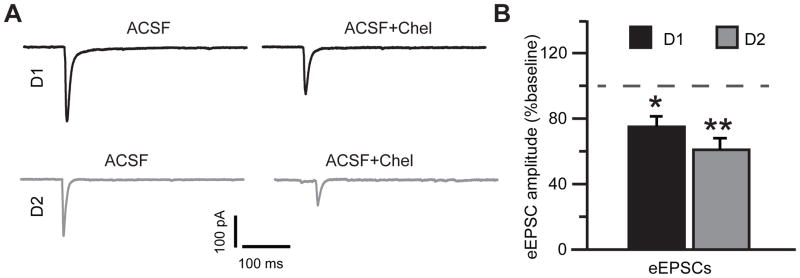
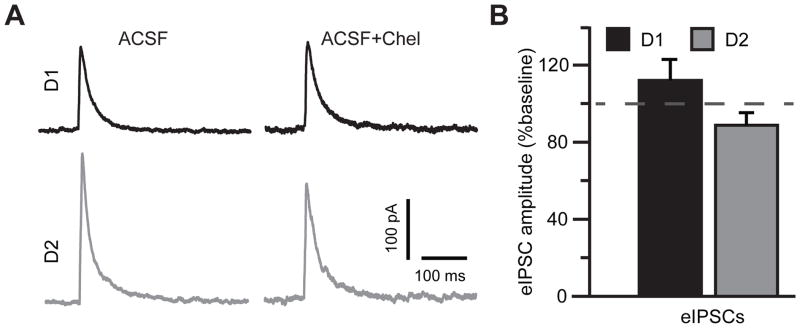
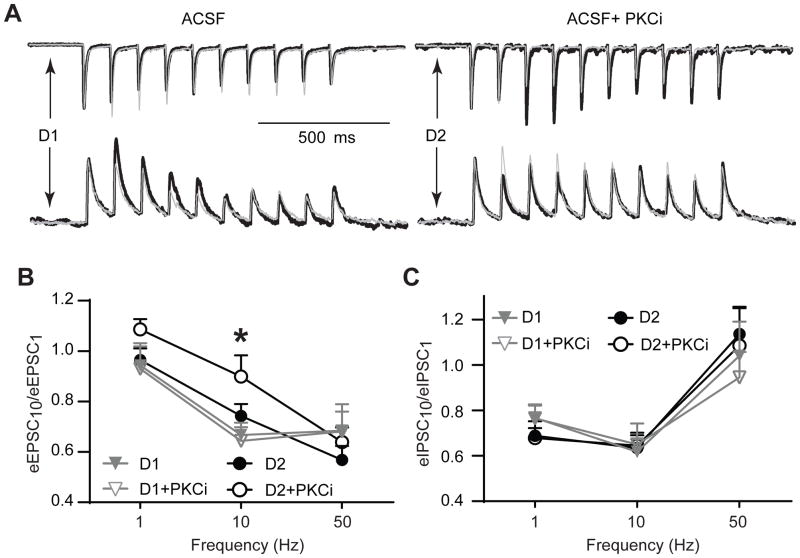
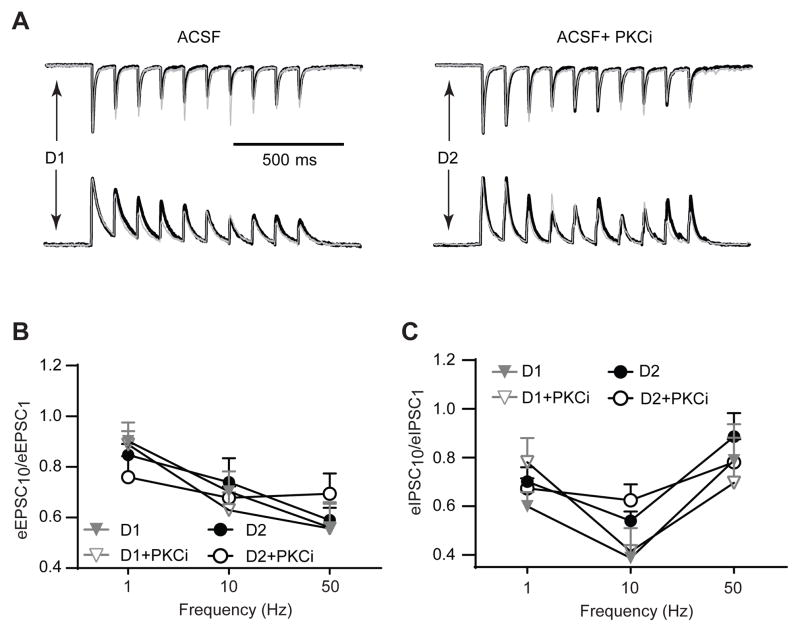
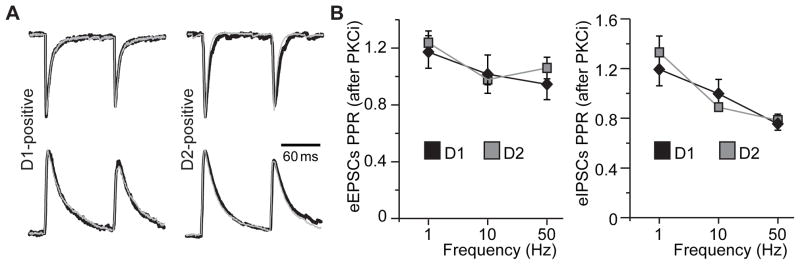
Stimulation of D1-like dopamine receptors (D1DRs) or D2-like dopamine receptors (D2DRs) in the nucleus accumbens (NAc) shell reinstates cocaine seeking in rats, an animal model of relapse. D2DRs and D1DRs activate protein kinase C (PKC) and recent studies indicate that activation of PKC in the NAc plays an important role in the reinstatement of drug seeking induced by a systemic cocaine priming injection. In the present study, pharmacological inhibition of PKC in the NAc shell attenuated cocaine seeking induced by intra-accumbens shell microinjection of a D2DR agonist, but not a D1DR agonist. D1DRs and D2DRs are primarily expressed on different accumbens medium spiny (MSN) neurons. Neuronal signaling and activity were assessed in these two populations of NAc neurons with transgenic mice expressing fluorescent labels under the control of D1DR and D2DR promoters. Following the extinction of cocaine self-administration, bath application of a PKC inhibitor produced similar effects on single evoked excitatory and inhibitory post-synaptic currents in D1DR- and D2DR-positive MSNs in the NAc shell. However, inhibition of PKC preferentially improved the ability of excitatory, but not inhibitory, synapses to sustain responding to brief train of stimuli specifically in D2DR-positive MSNs. This effect did not appear to involve modulation of presynaptic release mechanisms. Taken together, these findings indicate that the reinstatement of cocaine seeking is at least partially due to D2DR-dependent increases in PKC signaling in the NAc shell, which reduce excitatory synaptic efficacy in D2DR-expressing MSNs.
Keywords: Addiction; Dopamine; EPSCs; Electrophysiology; IPSCs; Relapse; Self-administration; Striatum.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Conflict of interest statement
The authors declare no potential conflict of interest relating to this study.
Figures
References
-
- Albin RL, Young AB, Penney JB. The functional anatomy of disorders of the basal ganglia. Trends Neurosci. 1995;18:63–64. - PubMed
-
- Anderson SM, Bari AA, Pierce RC. Administration of the D1-like dopamine receptor antagonist SCH-23390 into the medial nucleus accumbens shell attenuates cocaine priming-induced reinstatement of drug-seeking behavior in rats. Psychopharmacology (Berl) 2003;168:132–138. - PubMed
-
- Anderson SM, Famous KR, Sadri-Vakili G, Kumaresan V, Schmidt HD, Bass CE, Terwilliger EF, Cha JH, Pierce RC. CaMKII: a biochemical bridge linking accumbens dopamine and glutamate systems in cocaine seeking. Nat Neurosci. 2008;11:344–353. - PubMed
-
- Anderson SM, Pierce RC. Cocaine-induced alterations in dopamine receptor signaling: implications for reinforcement and reinstatement. Pharmacol Ther. 2005;106:389–403. - PubMed
-
- Anderson SM, Schmidt HD, Pierce RC. Administration of the D2 dopamine receptor antagonist sulpiride into the shell, but not the core, of the nucleus accumbens attenuates cocaine priming-induced reinstatement of drug seeking. Neuropsychopharmacology. 2006;31:1452–1461. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
