

Diverse mechanisms for spliceosome-mediated 3' end processing of telomerase RNA
- PMID: 25598145
- PMCID: PMC4299874
- DOI: 10.1038/ncomms7104
Diverse mechanisms for spliceosome-mediated 3' end processing of telomerase RNA
Abstract
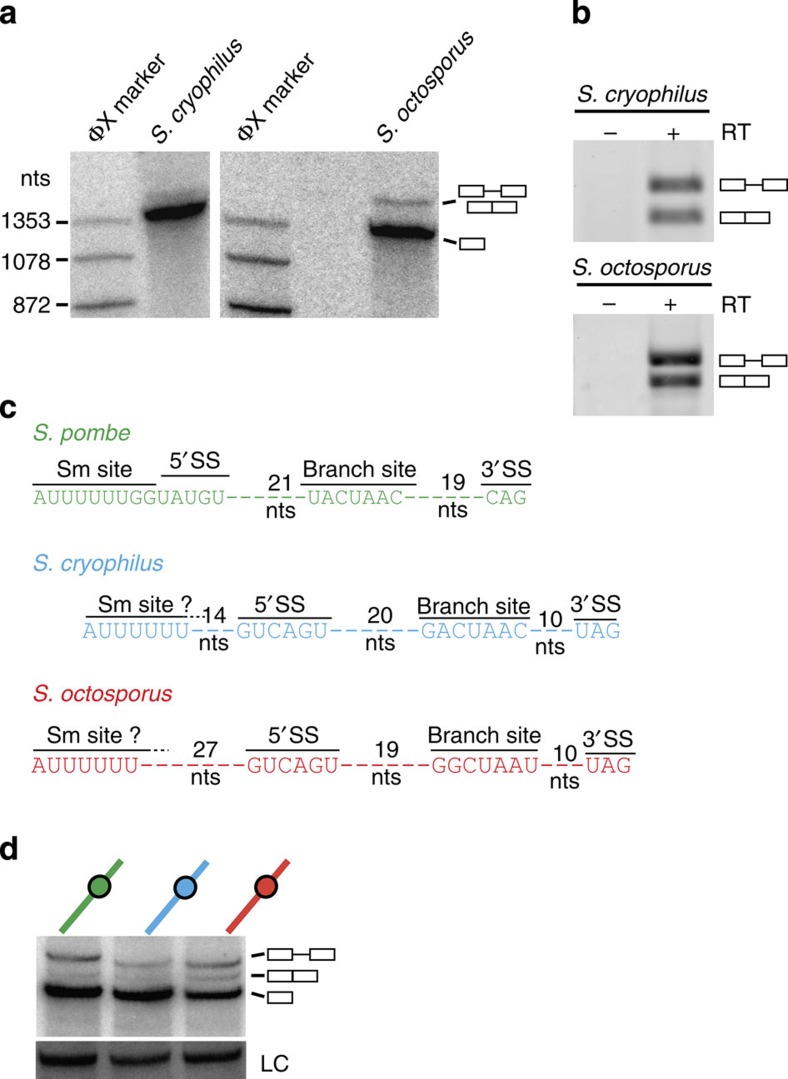
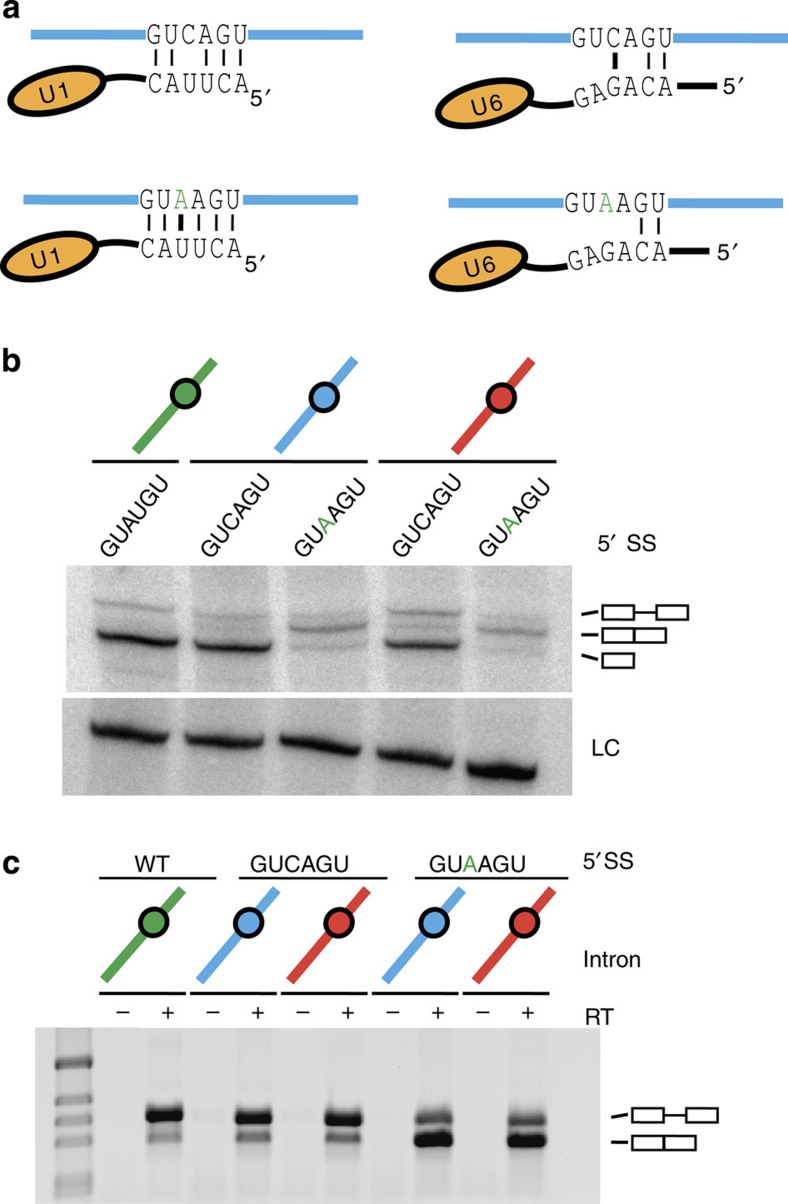
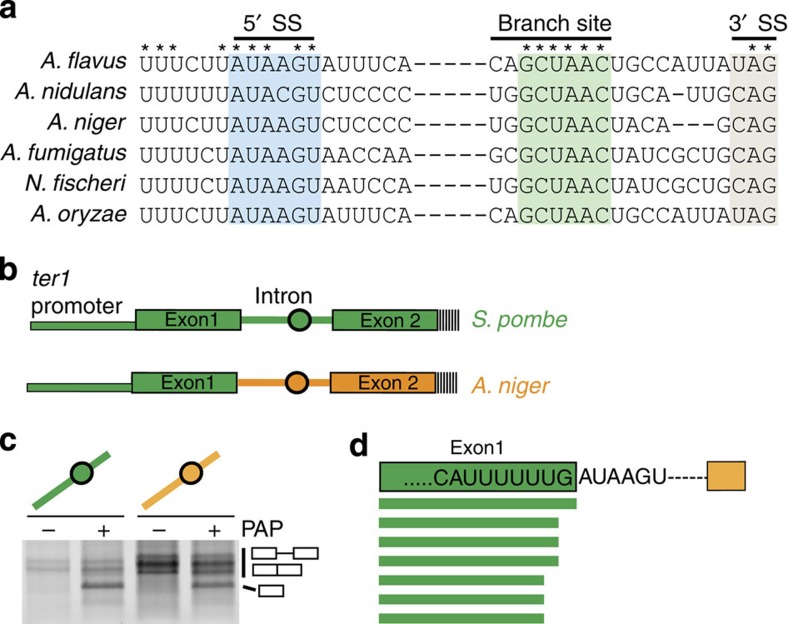
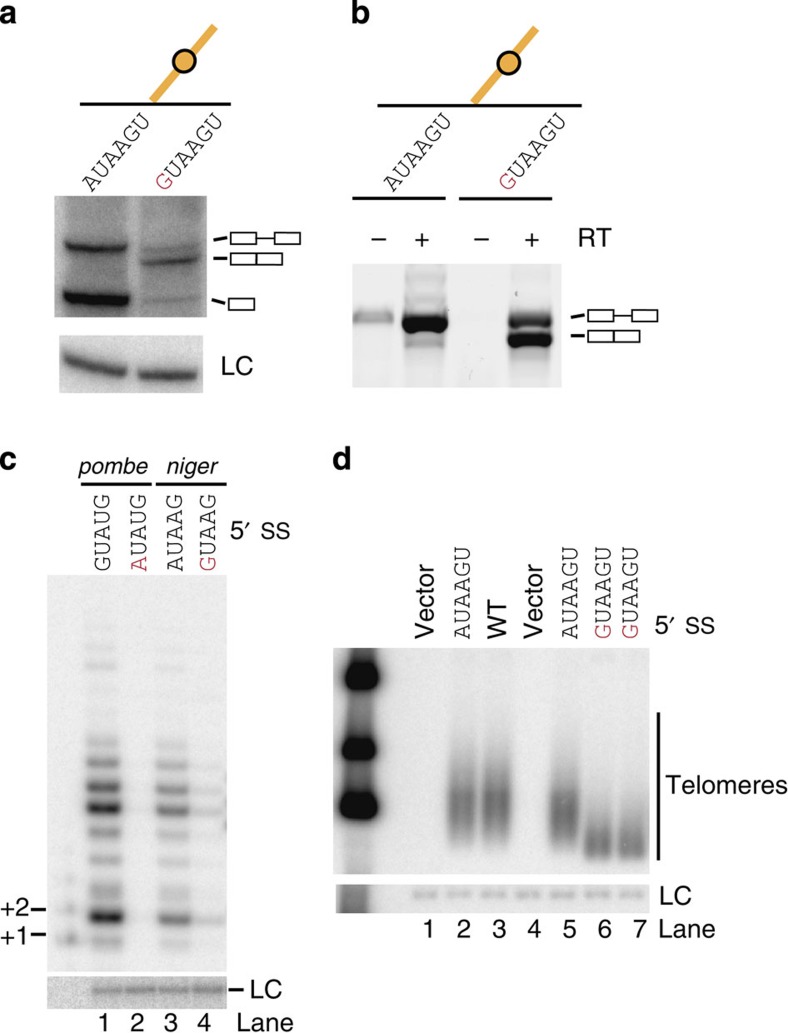
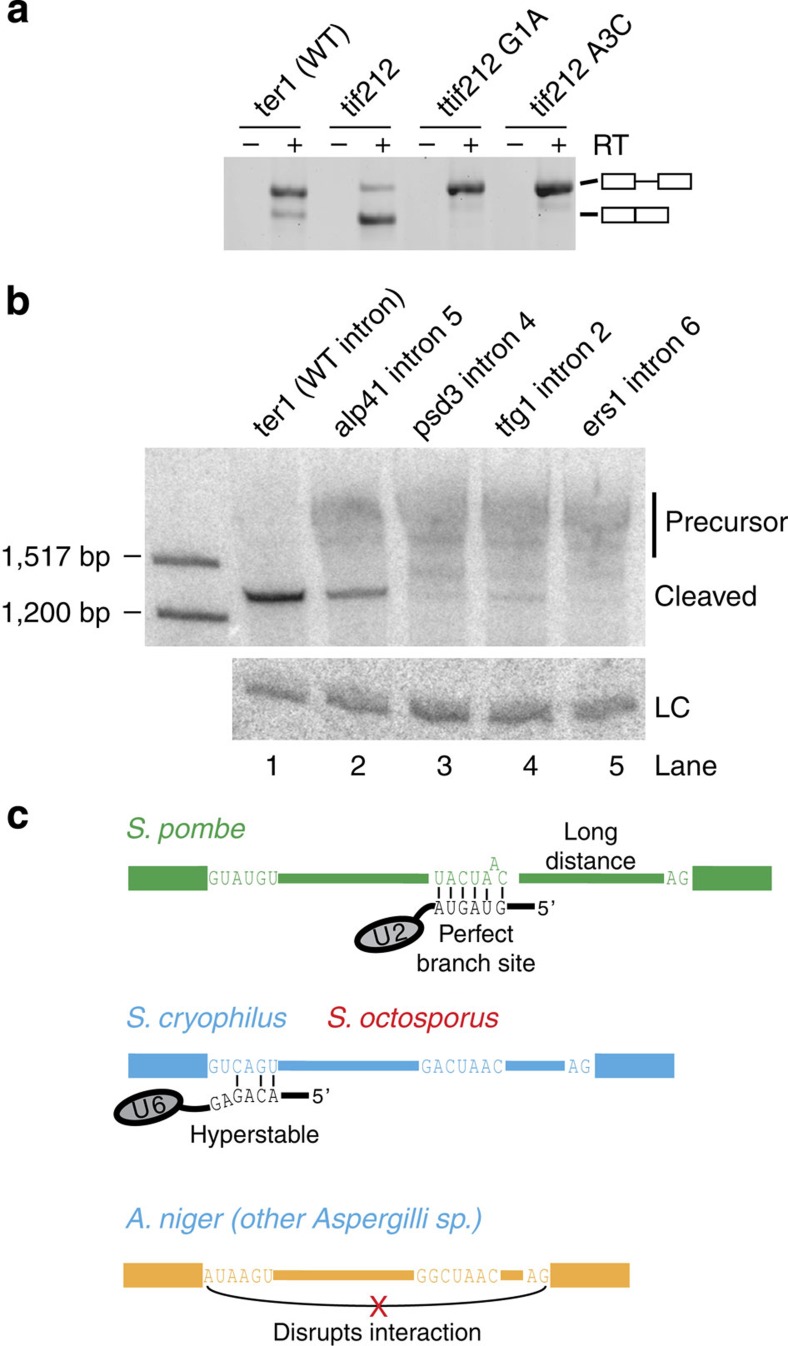
The 3' end of Schizosaccharomyces pombe telomerase RNA (SpTER1) is generated by spliceosomal cleavage, a reaction that corresponds to the first step of splicing. The observation that the spliceosome functions in 3' end processing raised questions about the evolutionary origin and conservation of this mechanism. We now present data in support of spliceosomes generating 3' ends of telomerase RNAs in other fungi. Strikingly, the mechanistic basis for restricting spliceosomal splicing to the first transesterification reaction differs substantially among species. Unlike S. pombe, two other fission yeasts rely on hyperstabilization of the U6 snRNA-5' splice site interaction to impede the 2nd step of splicing. In contrast, a non-canonical 5' splice site blocks the second transesterification reaction in Aspergillus species. These results demonstrate a conserved role for spliceosomes functioning in 3' end processing. Divergent mechanisms of uncoupling the two steps of splicing argue for multiple origins of this pathway.
Figures
Similar articles
-
Prevalent and distinct spliceosomal 3'-end processing mechanisms for fungal telomerase RNA.Nat Commun. 2015 Jan 19;6:6105. doi: 10.1038/ncomms7105. Nat Commun. 2015. PMID: 25598218 Free PMC article.
-
Spliceosomal cleavage generates the 3' end of telomerase RNA.Nature. 2008 Dec 18;456(7224):910-4. doi: 10.1038/nature07584. Epub 2008 Dec 3. Nature. 2008. PMID: 19052544
-
Intronic sequence elements impede exon ligation and trigger a discard pathway that yields functional telomerase RNA in fission yeast.Genes Dev. 2013 Mar 15;27(6):627-38. doi: 10.1101/gad.212738.112. Epub 2013 Mar 6. Genes Dev. 2013. PMID: 23468430 Free PMC article.
-
The evolutionary conservation of the splicing apparatus between fission yeast and man.Nucleic Acids Symp Ser. 1995;(33):226-8. Nucleic Acids Symp Ser. 1995. PMID: 8643378 Review.
-
An unexpected ending: noncanonical 3' end processing mechanisms.RNA. 2010 Feb;16(2):259-66. doi: 10.1261/rna.1907510. Epub 2009 Dec 9. RNA. 2010. PMID: 20007330 Free PMC article. Review.
Cited by
-
New perspectives on telomerase RNA structure and function.Wiley Interdiscip Rev RNA. 2018 Mar;9(2):10.1002/wrna.1456. doi: 10.1002/wrna.1456. Epub 2017 Nov 9. Wiley Interdiscip Rev RNA. 2018. PMID: 29124890 Free PMC article. Review.
-
A degenerate telomerase RNA directs telomeric DNA synthesis in lepidopteran insects.Proc Natl Acad Sci U S A. 2025 Mar 4;122(9):e2424443122. doi: 10.1073/pnas.2424443122. Epub 2025 Feb 28. Proc Natl Acad Sci U S A. 2025. PMID: 40020192 Free PMC article.
-
Peculiarities of Yeasts and Human Telomerase RNAs Processing.Acta Naturae. 2016 Oct-Dec;8(4):14-22. Acta Naturae. 2016. PMID: 28050263 Free PMC article.
-
Early splicing functions of fission yeast Prp16 and its unexpected requirement for gene Silencing is governed by intronic features.RNA Biol. 2019 Jun;16(6):754-769. doi: 10.1080/15476286.2019.1585737. Epub 2019 Mar 20. RNA Biol. 2019. PMID: 30810475 Free PMC article.
-
A high-quality reference genome for the fission yeast Schizosaccharomyces osmophilus.G3 (Bethesda). 2023 Apr 11;13(4):jkad028. doi: 10.1093/g3journal/jkad028. G3 (Bethesda). 2023. PMID: 36748990 Free PMC article.
References
-
- Nakamura T. M. et al.. Telomerase catalytic subunit homologs from fission yeast and human. Science 277, 955–959 (1997). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
