

B cell follicle sanctuary permits persistent productive simian immunodeficiency virus infection in elite controllers
- PMID: 25599132
- PMCID: PMC4320022
- DOI: 10.1038/nm.3781
B cell follicle sanctuary permits persistent productive simian immunodeficiency virus infection in elite controllers
Abstract
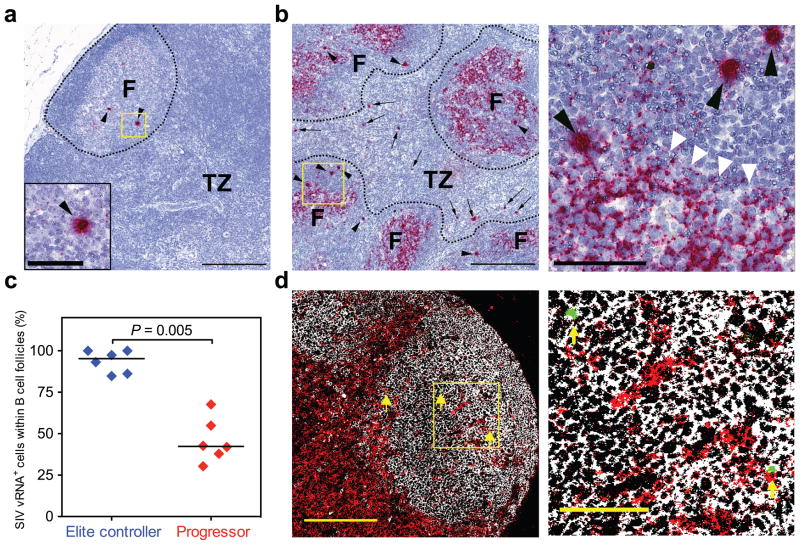
Chronic-phase HIV and simian immunodeficiency virus (SIV) replication is reduced by as much as 10,000-fold in elite controllers (ECs) compared with typical progressors (TPs), but sufficient viral replication persists in EC tissues to allow viral sequence evolution and induce excess immune activation. Here we show that productive SIV infection in rhesus monkey ECs, but not TPs, is markedly restricted to CD4(+) follicular helper T (TFH) cells, suggesting that these EC monkeys' highly effective SIV-specific CD8(+) T cells can effectively clear productive SIV infection from extrafollicular sites, but their relative exclusion from B cell follicles prevents their elimination of productively infected TFH cells. CD8(+) lymphocyte depletion in EC monkeys resulted in a dramatic re-distribution of productive SIV infection to non-TFH cells, with restriction of productive infection to TFH cells resuming upon CD8(+) T cell recovery. Thus, B cell follicles constitute 'sanctuaries' for persistent SIV replication in the presence of potent anti-viral CD8(+) T cell responses, potentially complicating efforts to cure HIV infection with therapeutic vaccination or T cell immunotherapy.
Conflict of interest statement
Figures





Comment in
-
AIDS virus seeks refuge in B cell follicles.Nat Med. 2015 Feb;21(2):111-2. doi: 10.1038/nm.3795. Nat Med. 2015. PMID: 25654598 No abstract available.
References
-
- Kirchhoff F. Immune evasion and counteraction of restriction factors by HIV-1 and other primate lentiviruses. Cell Host Microbe. 2010;8:55–67. - PubMed
-
- Deeks SG, Walker BD. Human immunodeficiency virus controllers: mechanisms of durable virus control in the absence of antiretroviral therapy. Immunity. 2007;27:406–416. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
