

Comparative RNA-Seq analysis reveals pervasive tissue-specific alternative polyadenylation in Caenorhabditis elegans intestine and muscles
- PMID: 25601023
- PMCID: PMC4343181
- DOI: 10.1186/s12915-015-0116-6
Comparative RNA-Seq analysis reveals pervasive tissue-specific alternative polyadenylation in Caenorhabditis elegans intestine and muscles
Abstract
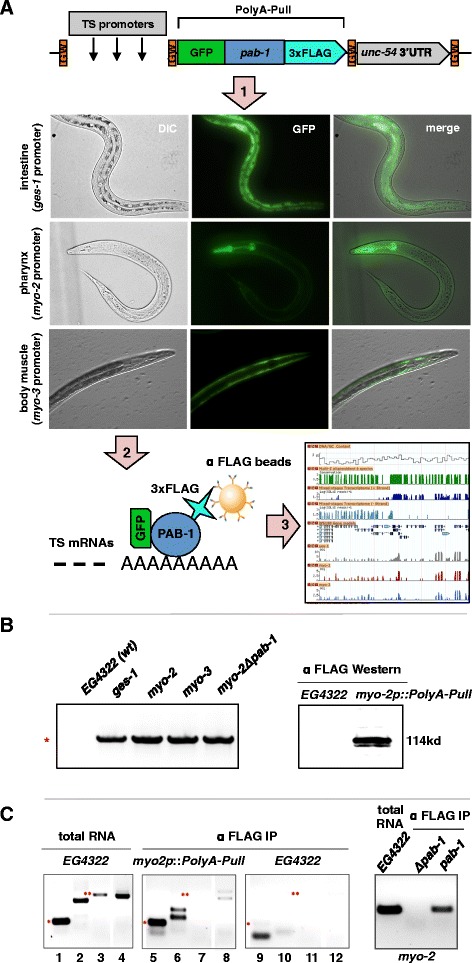
Background: Tissue-specific RNA plasticity broadly impacts the development, tissue identity and adaptability of all organisms, but changes in composition, expression levels and its impact on gene regulation in different somatic tissues are largely unknown. Here we developed a new method, polyA-tagging and sequencing (PAT-Seq) to isolate high-quality tissue-specific mRNA from Caenorhabditis elegans intestine, pharynx and body muscle tissues and study changes in their tissue-specific transcriptomes and 3'UTRomes.
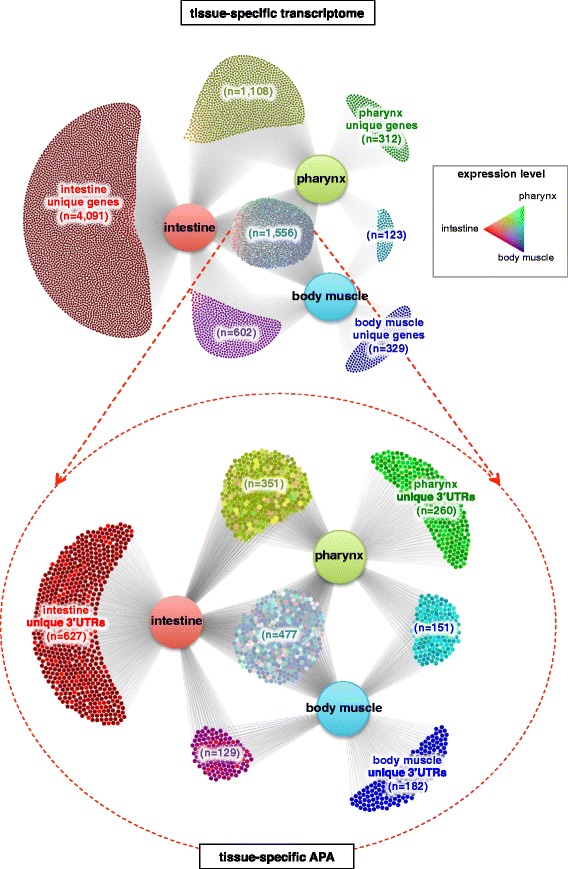
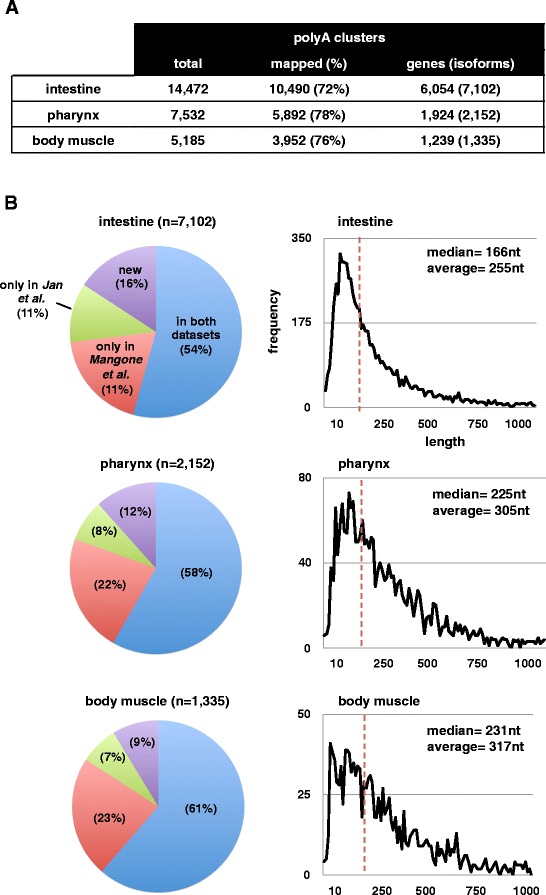
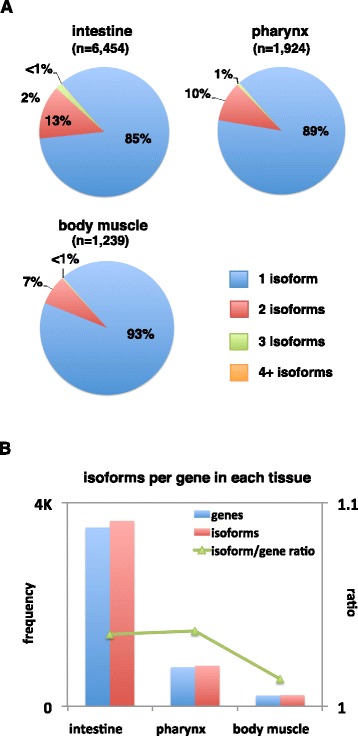
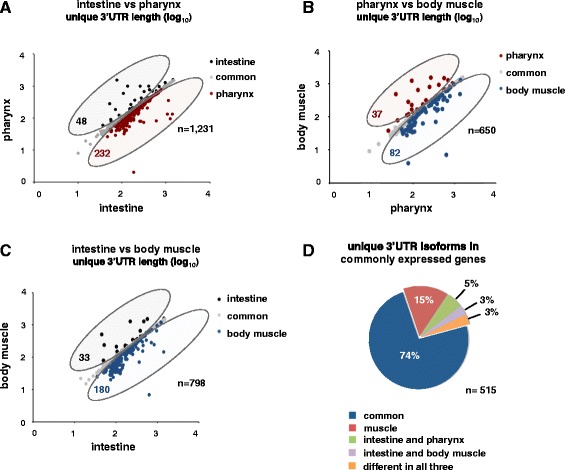
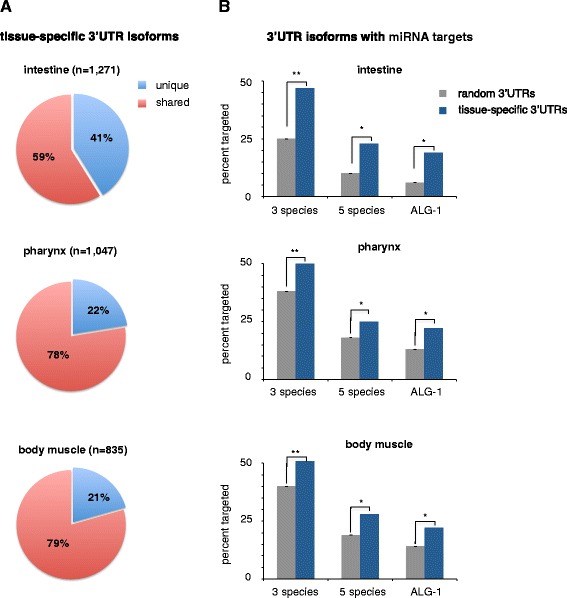
Results: We have identified thousands of novel genes and isoforms differentially expressed between these three tissues. The intestine transcriptome is expansive, expressing over 30% of C. elegans mRNAs, while muscle transcriptomes are smaller but contain characteristic unique gene signatures. Active promoter regions in all three tissues reveal both known and novel enriched tissue-specific elements, along with putative transcription factors, suggesting novel tissue-specific modes of transcription initiation. We have precisely mapped approximately 20,000 tissue-specific polyadenylation sites and discovered that about 30% of transcripts in somatic cells use alternative polyadenylation in a tissue-specific manner, with their 3'UTR isoforms significantly enriched with microRNA targets.
Conclusions: For the first time, PAT-Seq allowed us to directly study tissue specific gene expression changes in an in vivo setting and compare these changes between three somatic tissues from the same organism at single-base resolution within the same experiment. We pinpoint precise tissue-specific transcriptome rearrangements and for the first time link tissue-specific alternative polyadenylation to miRNA regulation, suggesting novel and unexplored tissue-specific post-transcriptional regulatory networks in somatic cells.
Figures
References
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
