

Oxidative stress conditions increase the frequency of de novo formation of the yeast [PSI+] prion
- PMID: 25601439
- PMCID: PMC4407919
- DOI: 10.1111/mmi.12930
Oxidative stress conditions increase the frequency of de novo formation of the yeast [PSI+] prion
Abstract
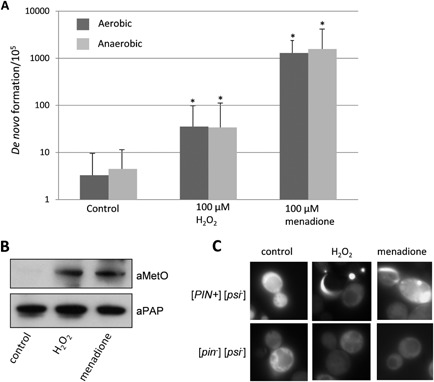
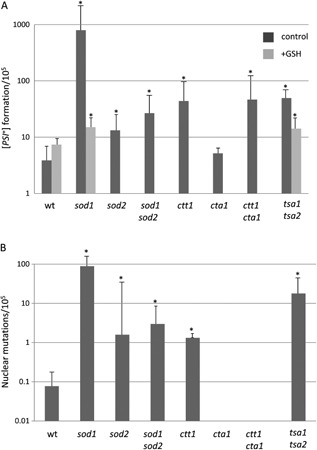
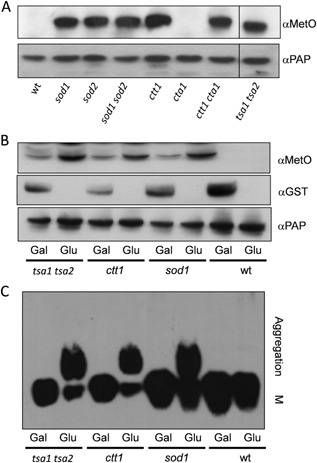
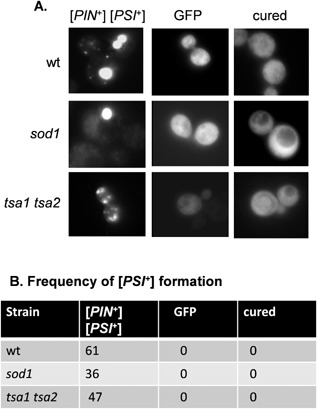
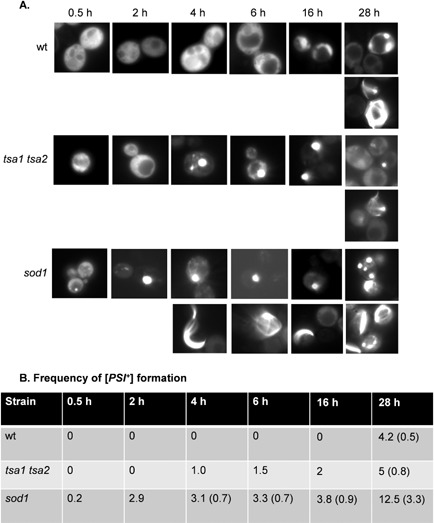
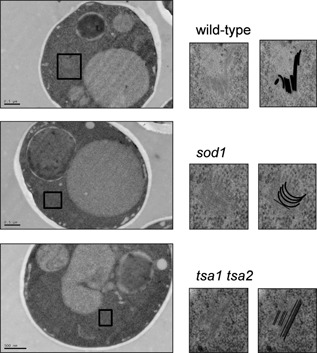
Prions are self-perpetuating amyloid protein aggregates which underlie various neurodegenerative diseases in mammals and heritable traits in yeast. The molecular basis of how yeast and mammalian prions form spontaneously into infectious amyloid-like structures is poorly understood. We have explored the hypothesis that oxidative stress is a general trigger for prion formation using the yeast [PSI(+)] prion, which is the altered conformation of the Sup35 translation termination factor. We show that the frequency of [PSI(+)] prion formation is elevated under conditions of oxidative stress and in mutants lacking key antioxidants. We detect increased oxidation of Sup35 methionine residues in antioxidant mutants and show that overexpression of methionine sulphoxide reductase abrogates both the oxidation of Sup35 and its conversion to the [PSI(+)] prion. [PSI(+)] prion formation is particularly elevated in a mutant lacking the Sod1 Cu,Zn-superoxide dismutase. We have used fluorescence microscopy to show that the de novo appearance of [PSI(+)] is both rapid and increased in frequency in this mutant. Finally, electron microscopy analysis of native Sup35 reveals that similar fibrillar structures are formed in both the wild-type and antioxidant mutants. Together, our data indicate that oxidative stress is a general trigger of [PSI(+) formation, which can be alleviated by antioxidant defenses.
© 2015 The Authors. Molecular Microbiology published by John Wiley & Sons Ltd.
Figures
References
-
- Alberti, S. , Halfmann, R. , and Lindquist, S. (2010) Biochemical, cell biological, and genetic assays to analyze amyloid and prion aggregation in yeast. Methods Enzymol 470: 709–734. - PubMed
-
- Berlett, B.S. , and Stadtman, E.R. (1997) Protein oxidation in aging, disease, and oxidative stress. J Biol Chem 272: 20313–20316. - PubMed
-
- Bradley, M.E. , and Liebman, S.W. (2004) The Sup35 domains required for maintenance of weak, strong or undifferentiated yeast [PSI+] prions. Mol Microbiol 51: 1649–1659. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
