

WT1 recruits TET2 to regulate its target gene expression and suppress leukemia cell proliferation
- PMID: 25601757
- PMCID: PMC4336627
- DOI: 10.1016/j.molcel.2014.12.023
WT1 recruits TET2 to regulate its target gene expression and suppress leukemia cell proliferation
Abstract
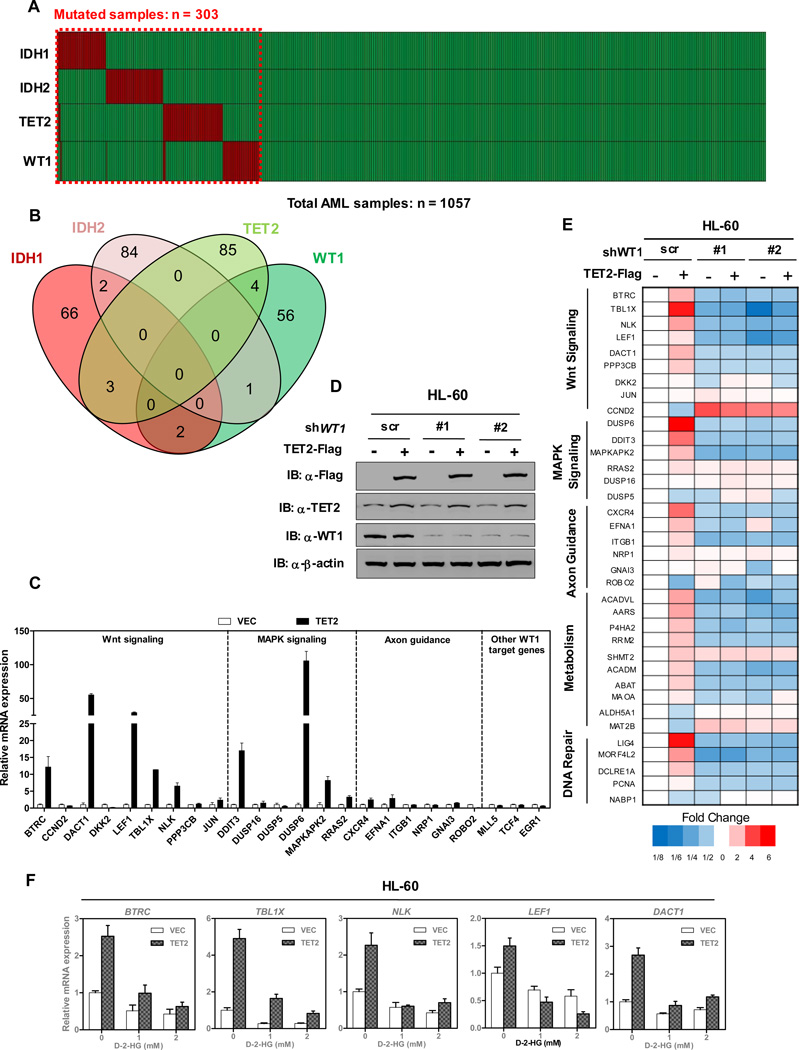
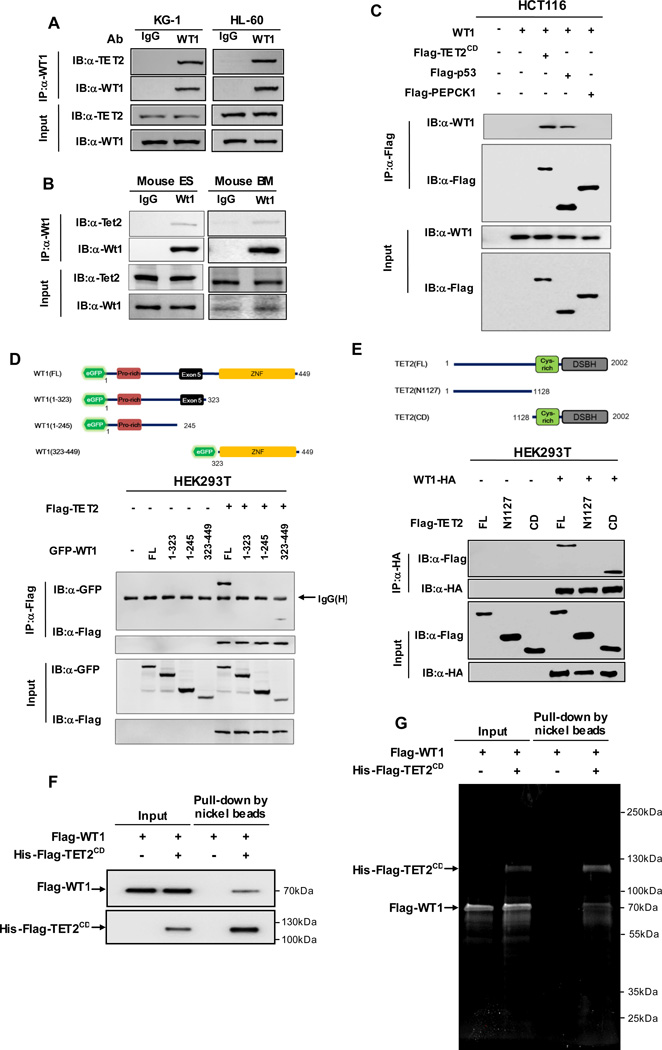
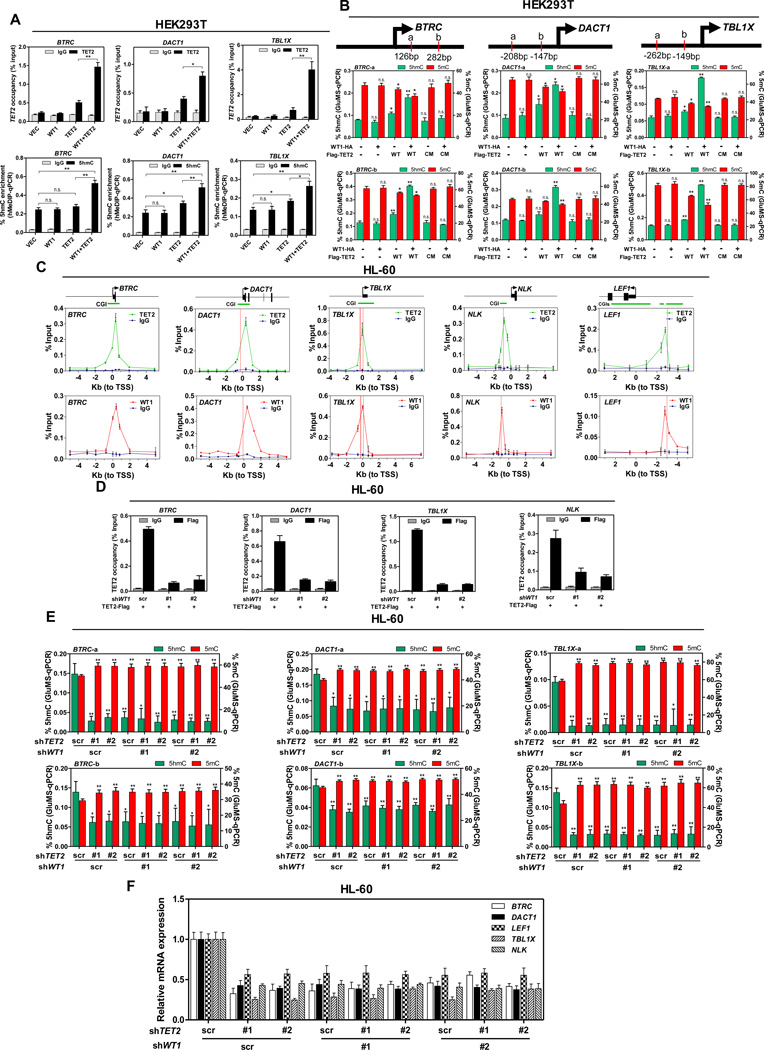
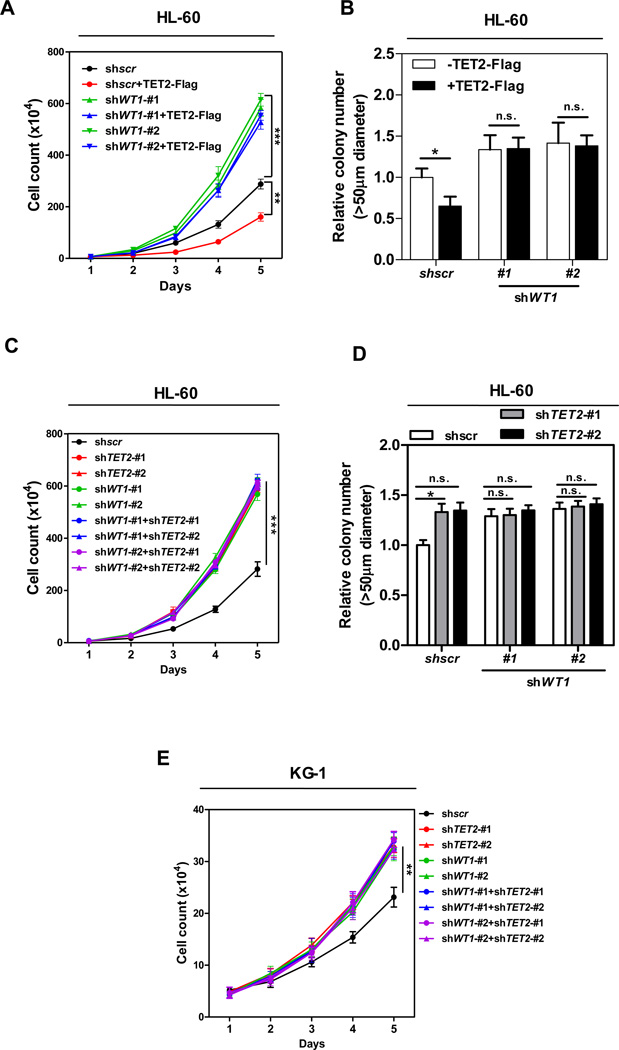
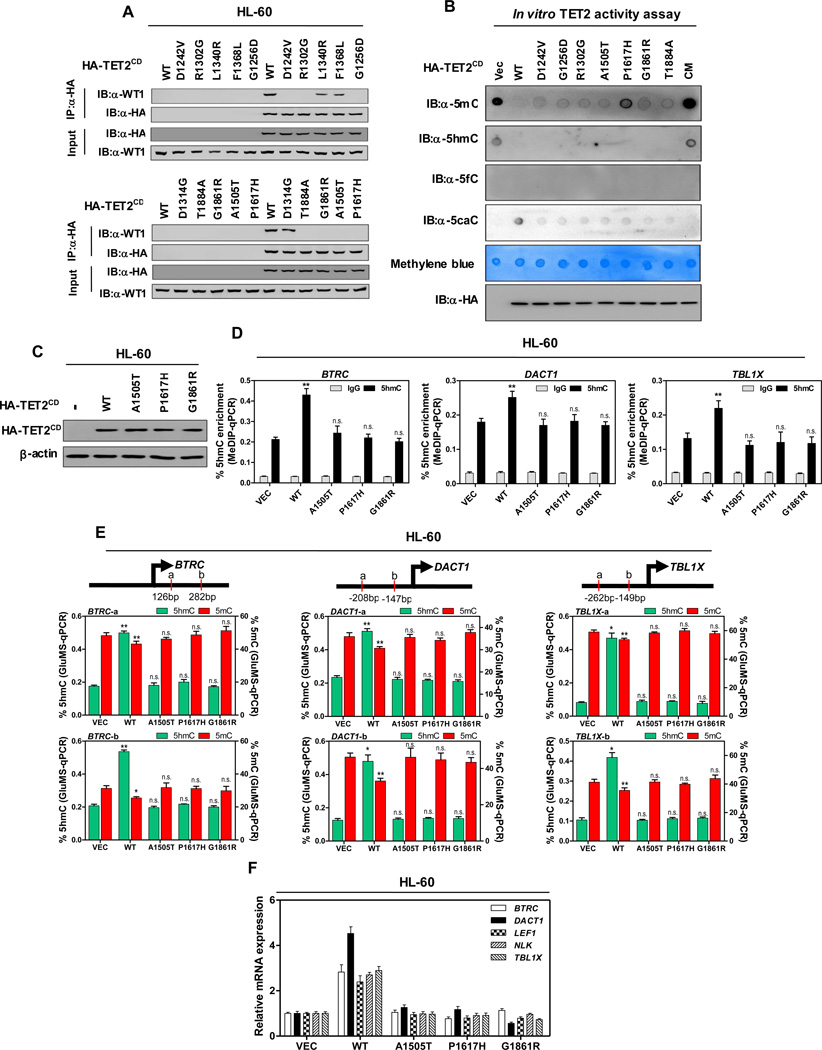
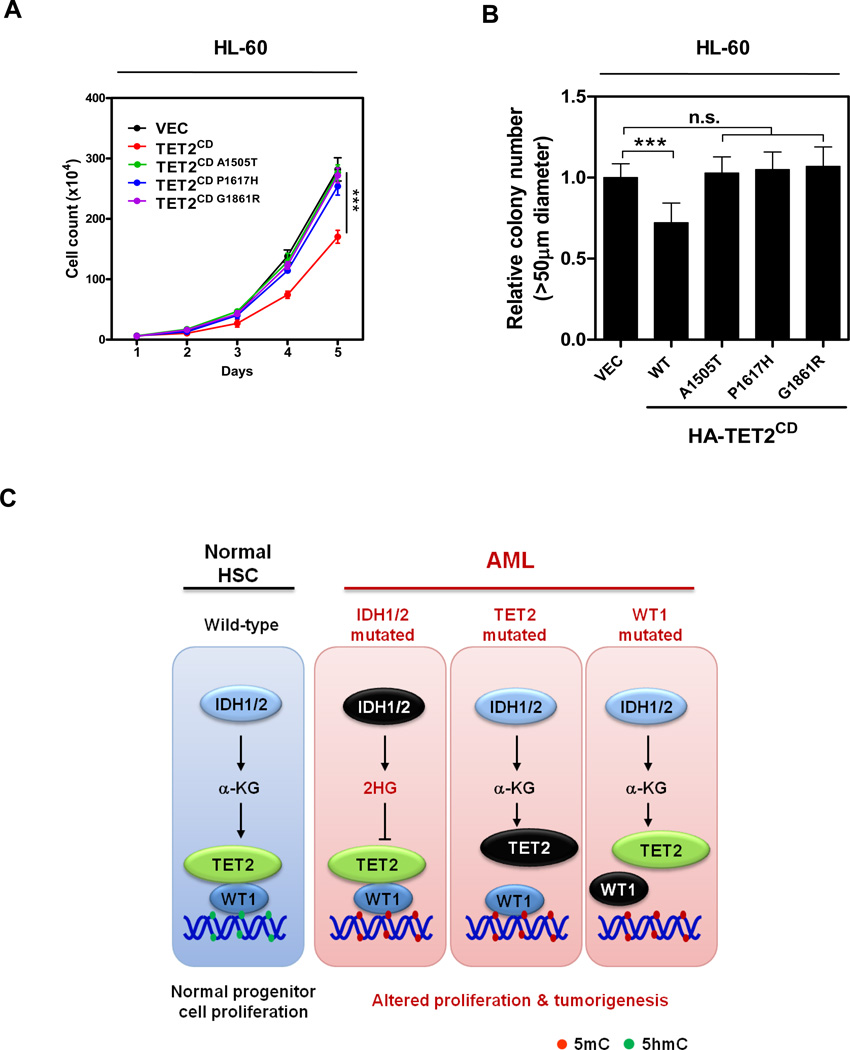
The TET2 DNA dioxygenase regulates cell identity and suppresses tumorigenesis by modulating DNA methylation and expression of a large number of genes. How TET2, like most other chromatin-modifying enzymes, is recruited to specific genomic sites is unknown. Here we report that WT1, a sequence-specific transcription factor, is mutated in a mutually exclusive manner with TET2, IDH1, and IDH2 in acute myeloid leukemia (AML). WT1 physically interacts with and recruits TET2 to its target genes to activate their expression. The interaction between WT1 and TET2 is disrupted by multiple AML-derived TET2 mutations. TET2 suppresses leukemia cell proliferation and colony formation in a manner dependent on WT1. These results provide a mechanism for targeting TET2 to a specific DNA sequence in the genome. Our results also provide an explanation for the mutual exclusivity of WT1 and TET2 mutations in AML, and suggest an IDH1/2-TET2-WT1 pathway in suppressing AML.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
Comment in
-
A new path to leukemia with WIT.Mol Cell. 2015 Feb 19;57(4):573-574. doi: 10.1016/j.molcel.2015.02.005. Mol Cell. 2015. PMID: 25699704
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
