

Endogenous cross-talk of fungal metabolites
- PMID: 25601857
- PMCID: PMC4283610
- DOI: 10.3389/fmicb.2014.00732
Endogenous cross-talk of fungal metabolites
Abstract
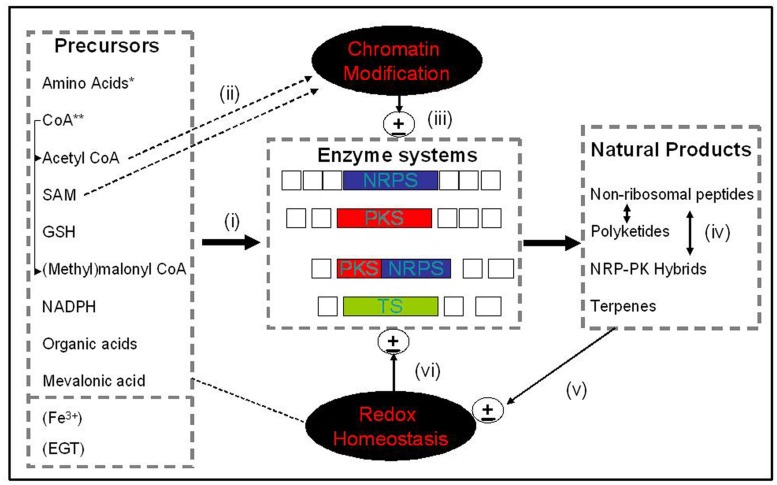
Non-ribosomal peptide (NRP) synthesis in fungi requires a ready supply of proteogenic and non-proteogenic amino acids which are subsequently incorporated into the nascent NRP via a thiotemplate mechanism catalyzed by NRP synthetases. Substrate amino acids can be modified prior to or during incorporation into the NRP, or following incorporation into an early stage amino acid-containing biosynthetic intermediate. These post-incorporation modifications involve a range of additional enzymatic activities including but not exclusively, monooxygenases, methyltransferases, epimerases, oxidoreductases, and glutathione S-transferases which are essential to effect biosynthesis of the final NRP. Likewise, polyketide biosynthesis is directly by polyketide synthase megaenzymes and cluster-encoded ancillary decorating enzymes. Additionally, a suite of additional primary metabolites, for example: coenzyme A (CoA), acetyl CoA, S-adenosylmethionine, glutathione (GSH), NADPH, malonyl CoA, and molecular oxygen, amongst others are required for NRP and polyketide synthesis (PKS). Clearly these processes must involve exquisite orchestration to facilitate the simultaneous biosynthesis of different types of NRPs, polyketides, and related metabolites requiring identical or similar biosynthetic precursors or co-factors. Moreover, the near identical structures of many natural products within a given family (e.g., ergot alkaloids), along with localization to similar regions within fungi (e.g., conidia) suggests that cross-talk may exist, in terms of biosynthesis and functionality. Finally, we speculate if certain biosynthetic steps involved in NRP and PKS play a role in cellular protection or environmental adaptation, and wonder if these enzymatic reactions are of equivalent importance to the actual biosynthesis of the final metabolite.
Keywords: NRPS; PKS; gliotoxin; natural products; redox homeostasis; secondary metabolites; siderophores; systems biology.
Figures
Similar articles
-
Nonribosomal peptide synthesis in Aspergillus fumigatus and other fungi.Microbiology (Reading). 2007 May;153(Pt 5):1297-1306. doi: 10.1099/mic.0.2006/006908-0. Microbiology (Reading). 2007. PMID: 17464044 Review.
-
Enzymes involved in fatty acid and polyketide biosynthesis in Streptomyces glaucescens: role of FabH and FabD and their acyl carrier protein specificity.Biochemistry. 2002 Aug 20;41(33):10462-71. doi: 10.1021/bi0258804. Biochemistry. 2002. PMID: 12173933
-
Novel features in a combined polyketide synthase/non-ribosomal peptide synthetase: the myxalamid biosynthetic gene cluster of the myxobacterium Stigmatella aurantiaca Sga15.Chem Biol. 2001 Jan;8(1):59-69. doi: 10.1016/s1074-5521(00)00056-9. Chem Biol. 2001. PMID: 11182319
-
Biosynthesis of hybrid peptide-polyketide natural products.Curr Opin Drug Discov Devel. 2001 Mar;4(2):215-28. Curr Opin Drug Discov Devel. 2001. PMID: 11378961 Review.
-
Epothilone biosynthesis: assembly of the methylthiazolylcarboxy starter unit on the EpoB subunit.Chem Biol. 2001 Sep;8(9):899-912. doi: 10.1016/s1074-5521(01)00064-3. Chem Biol. 2001. PMID: 11564558
Cited by
-
How to improve the production of peptidyl compounds in filamentous fungi.Front Fungal Biol. 2022 Dec 22;3:1085624. doi: 10.3389/ffunb.2022.1085624. eCollection 2022. Front Fungal Biol. 2022. PMID: 37746201 Free PMC article.
-
Study of in vitro interaction between Fusarium verticillioides and Streptomyces sp. using metabolomics.Folia Microbiol (Praha). 2020 Apr;65(2):303-314. doi: 10.1007/s12223-019-00725-z. Epub 2019 Jun 28. Folia Microbiol (Praha). 2020. PMID: 31250362
-
Metabolites Identified during Varied Doses of Aspergillus Species in Zea mays Grains, and Their Correlation with Aflatoxin Levels.Toxins (Basel). 2018 May 7;10(5):187. doi: 10.3390/toxins10050187. Toxins (Basel). 2018. PMID: 29735944 Free PMC article.
-
Multiplex Base-Editing Enables Combinatorial Epigenetic Regulation for Genome Mining of Fungal Natural Products.J Am Chem Soc. 2023 Jan 11;145(1):413-421. doi: 10.1021/jacs.2c10211. Epub 2022 Dec 21. J Am Chem Soc. 2023. PMID: 36542862 Free PMC article.
-
Twists and Turns in the Salicylate Catabolism of Aspergillus terreus, Revealing New Roles of the 3-Hydroxyanthranilate Pathway.mSystems. 2021 Jan 26;6(1):e00230-20. doi: 10.1128/mSystems.00230-20. mSystems. 2021. PMID: 33500329 Free PMC article.
References
-
- Balibar C. J., Walsh C. T. (2006). GliP, a multimodular nonribosomal peptide synthetase in Aspergillus fumigatus, makes the diketopiperazine scaffold of gliotoxin. Biochemistry 45 15029–15038. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous