

Regulation of bacterial virulence gene expression by cell envelope stress responses
- PMID: 25603429
- PMCID: PMC4601401
- DOI: 10.4161/21505594.2014.965580
Regulation of bacterial virulence gene expression by cell envelope stress responses
Abstract
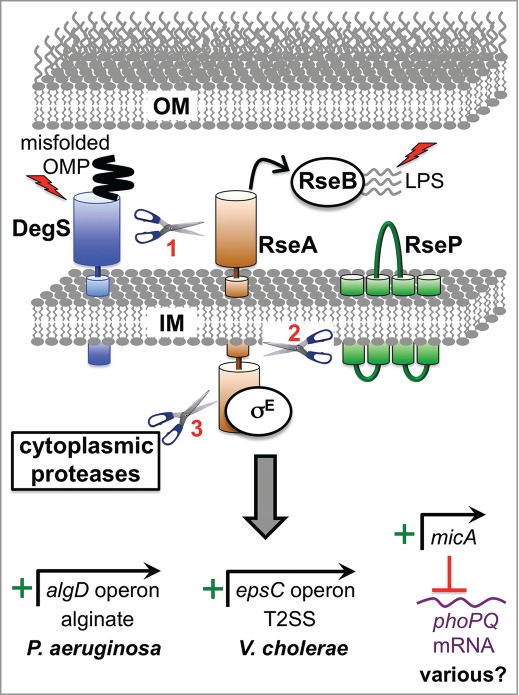
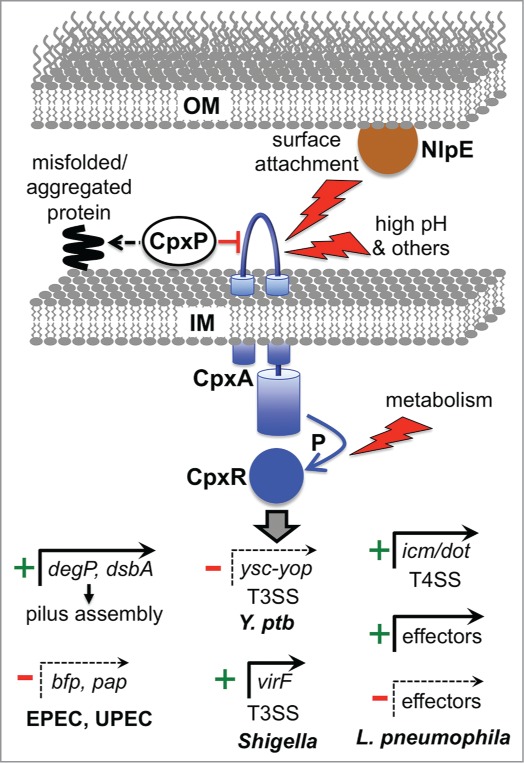
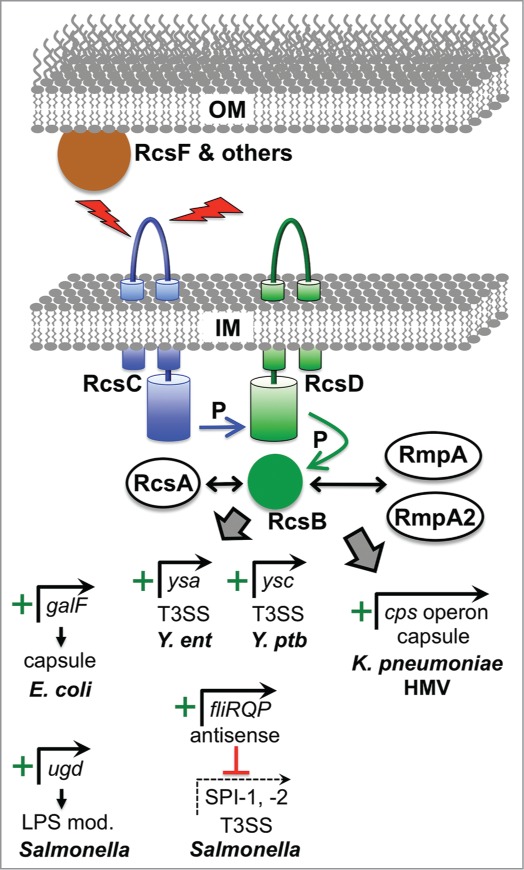
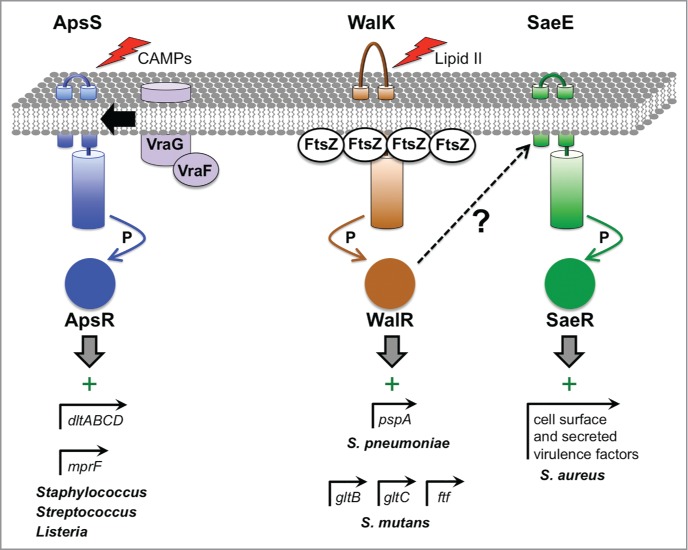
The bacterial cytoplasm lies within a multilayered envelope that must be protected from internal and external hazards. This protection is provided by cell envelope stress responses (ESRs), which detect threats and reprogram gene expression to ensure survival. Pathogens frequently need these ESRs to survive inside the host, where their envelopes face dangerous environmental changes and attack from antimicrobial molecules. In addition, some virulence genes have become integrated into ESR regulons. This might be because these genes can protect the cell envelope from damage by host molecules, or it might help ESRs to reduce stress by moderating the assembly of virulence factors within the envelope. Alternatively, it could simply be a mechanism to coordinate the induction of virulence gene expression with entry into the host. Here, we briefly describe some of the bacterial ESRs, followed by examples where they control virulence gene expression in both Gram-negative and Gram-positive pathogens.
Keywords: BFP, bundle-forming pilus; CAMP, cationic antimicrobial peptide; CF, cystic fibrosis; ECF, extracytoplasmic function; EPEC, enteropathogenic E. coli; ESR, envelope stress response; HMV, hypermucoviscosity; IM, inner membrane; LPS, lipopolysaccharide; LTA, lipoteichoic acids; OM, outer membrane; OMP, outer membrane protein; PG, phosphatidylglycerol; T(2/3/4)SS, type II/III/IV secretion system; UPEC, uropathogenic E. coli; WTA, wall teichoic acids; antimicrobial peptide; bacterial pathogens; cell envelope; gene regulation; peptidoglycan; phospholipid; stress response; teichoic acid; virulence gene.
Figures
References
-
- Raivio TL. Envelope stress responses and Gram-negative bacterial pathogenesis. Mol Microbiol 2005; 56:1119-28; PMID:15882407; http://dx.doi.org/ 10.1111/j.1365-2958.2005.04625.x - DOI - PubMed
-
- Rowley G, Spector M, Kormanec J, Roberts M. Pushing the envelope: extracytoplasmic stress responses in bacterial pathogens. Nat Rev Microbiol 2006; 4:383-94; PMID: 16715050; http://dx.doi.org/ 10.1038/nrmicro1394 - DOI - PubMed
-
- Silhavy TJ, Kahne D, Walker S. The bacterial cell envelope. Cold Spring Harb Perspect Biol 2010; 2:a000414; PMID:20452953; http://dx.doi.org/ 10.1101/cshperspect.a000414 - DOI - PMC - PubMed
-
- Raivio TL, Silhavy TJ. The sigmaE and Cpx regulatory pathways: overlapping but distinct envelope stress responses. Curr Opin Microbiol 1999; 2:159-65; PMID:10322173; http://dx.doi.org/ 10.1016/S1369-5274(99)80028-9 - DOI - PubMed
-
- Mecsas J, Rouviere PE, Erickson JW, Donohue TJ, Gross CA. The activity of sigma E, an Escherichia coli heat-inducible sigma-factor, is modulated by expression of outer membrane proteins. Genes Dev 1993; 7:2618-28; PMID:8276244; http://dx.doi.org/ 10.1101/gad.7.12b.2618 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous