

An association of Aquaporin-4 with the immunoregulation of liver pathology in mice infected with Schistosoma japonicum
- PMID: 25604731
- PMCID: PMC4311472
- DOI: 10.1186/s13071-015-0650-7
An association of Aquaporin-4 with the immunoregulation of liver pathology in mice infected with Schistosoma japonicum
Abstract
Background: Schistosomiasis is a chronic parasitic disease that affects approximately 200 million people. In Schistosomiasis japonica and mansoni, parasite eggs were trapped in host liver and stimulated the CD4(+)T cell responses to regulate the formation of the granulomas. Subsequently, excessive granulomatous response in some heavily, and/or repeatedly infected individuals could result in chronic liver fibrosis and circulatory impairment. Thus, elucidation of the mechanisms of these responses will not only provide more information to better understand the mechanisms of the immunoregulation in schistosomiasis, but also help to design new therapies to control granuloma-associated immunopathology. The role of aquaporin-4 (AQP4) in water transport has been extensively investigated in the central nervous system (CNS). Recently, studies have shown that AQP4 expresses in immune system and lack of AQP4 in mice results in significantly less CD4(+)CD25(+) T regulatory cells (Treg cells) under physiological condition, one of the subpopulations of CD4(+)T cells which restrains immunopathology in hosts with schistosomiasis. However, little information exists regarding the contribution of AQP4 to the immune regulation in schistosome infection.
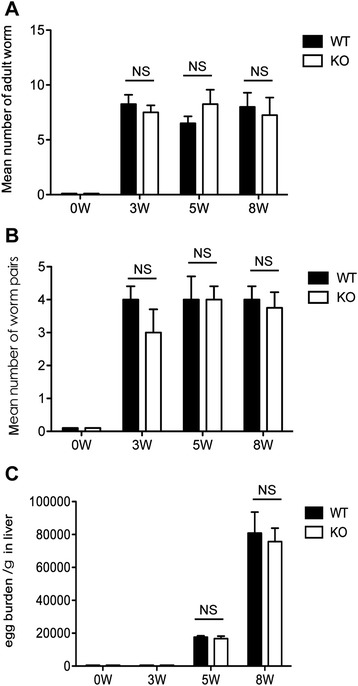
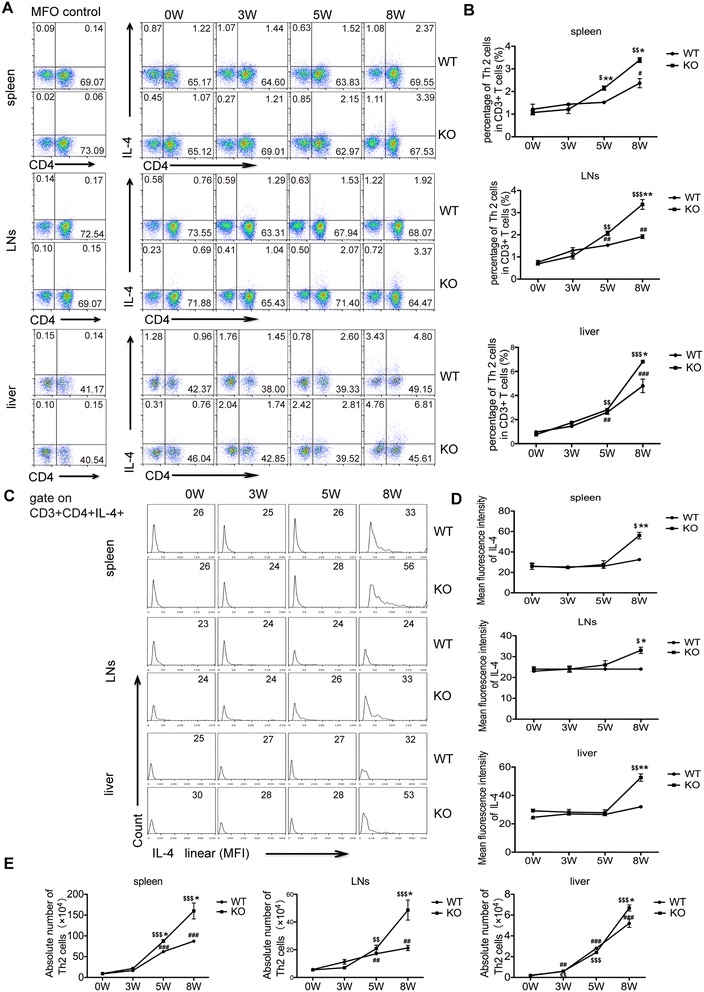
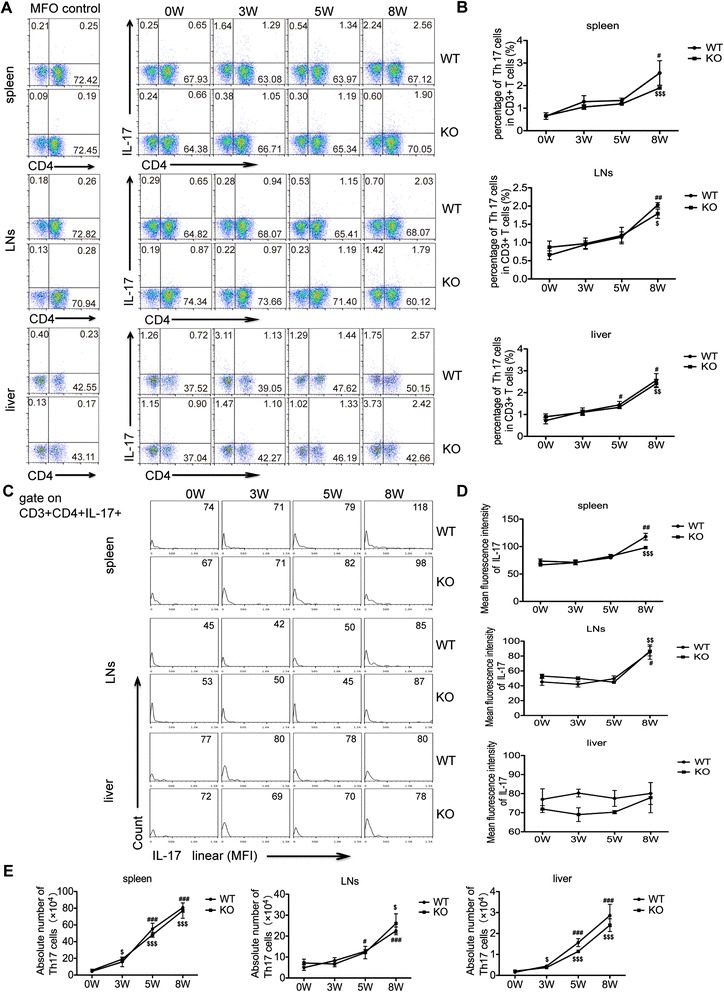
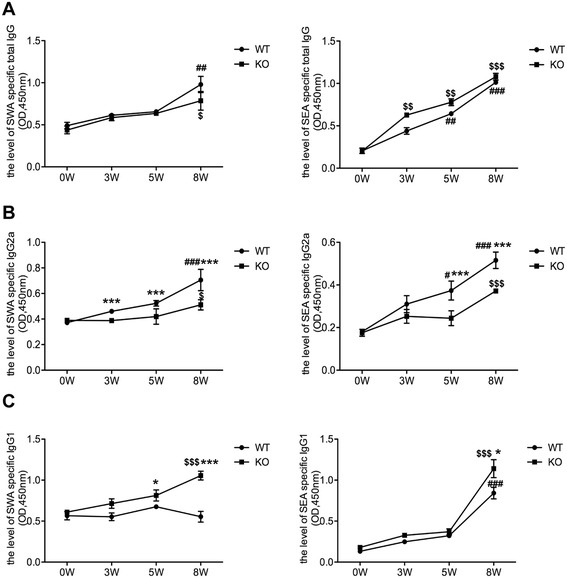
Methods: The liver granulomatous response in S. japonicum-infected AQP4 knockout (KO) mice and its wild-type (WT) littermates were detected by staining liver sections with hematoxylin and eosin. The generation of various CD4(+) T subsets, including Th1, Th2, Th17, and Treg cells were analyzed by flow cytometry. In addition, the levels of total IgG, IgG1, IgG2a in serum of infected mice were detected by ELISA assay.
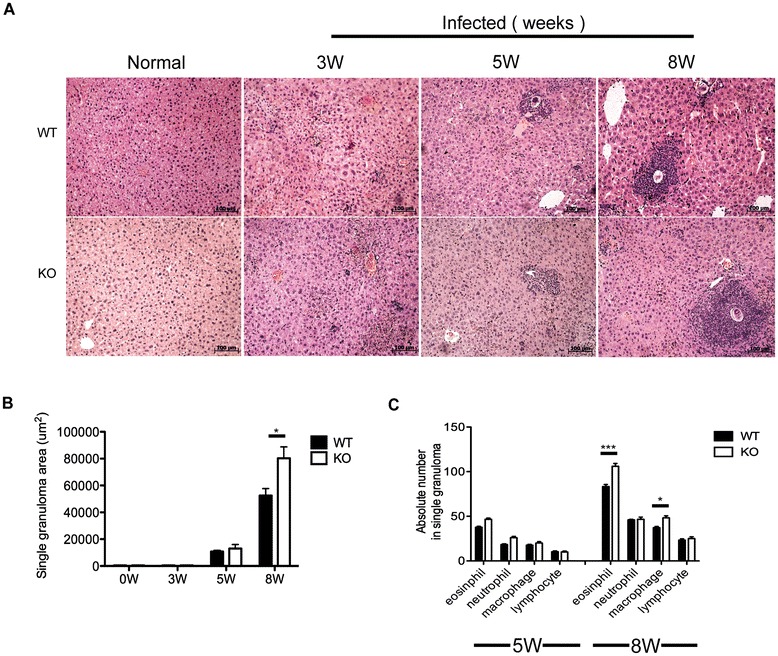
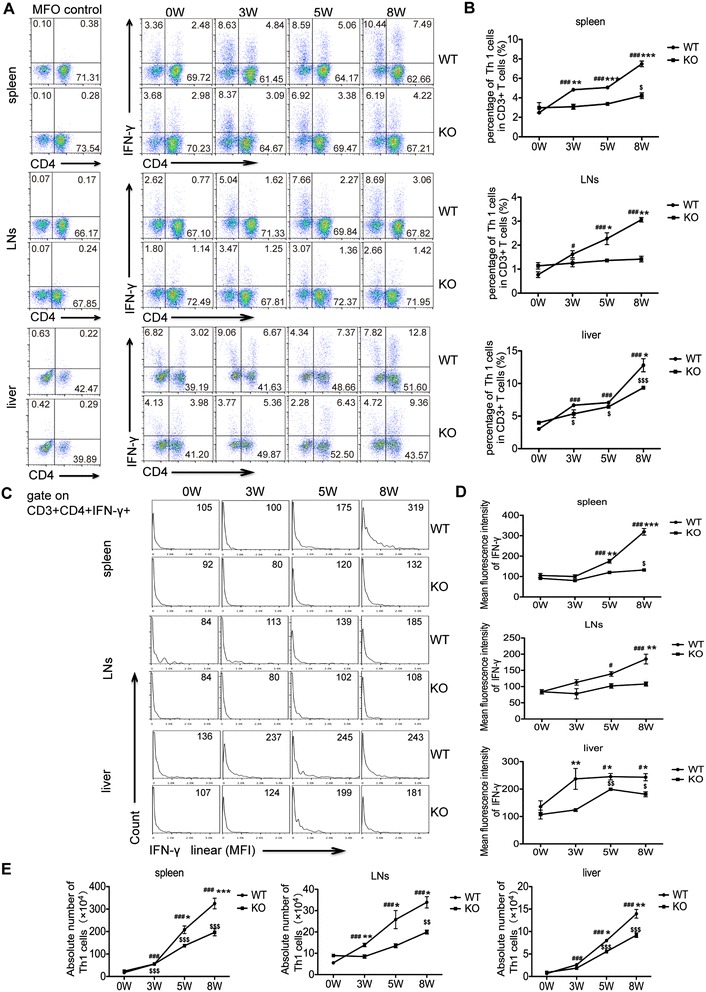
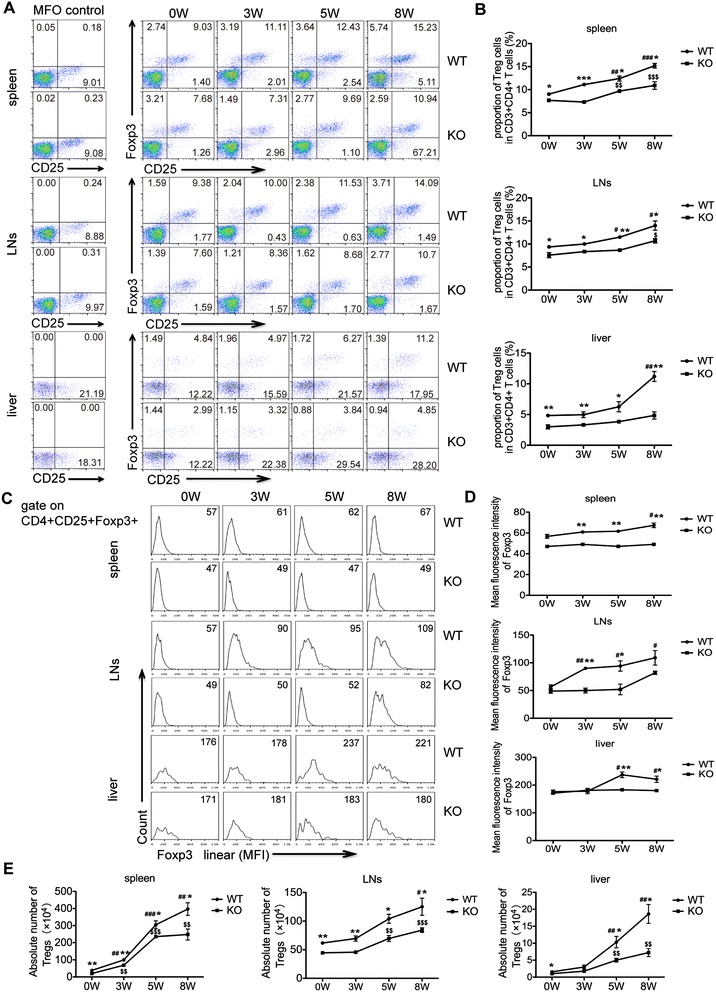
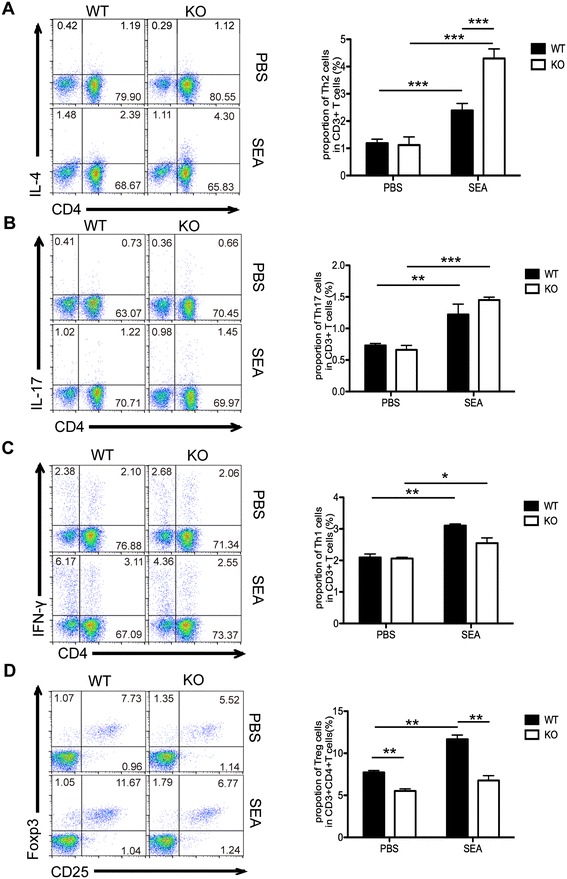
Results: Our results showed an enhanced granulomatous response with increased accumulation of eosinophils and macrophages around eggs in the liver of AQP4 KO mice with Schistosomiasis japonica. In addition, our study demonstrated enhanced Th2 but reduced Th1 and Treg cells generation in AQP4 KO mice with Schistosomiasis japonica, which may, at least partly, account for the enhancement of the liver granuloma formation.
Conclusion: Our study for the first time provides evidences that AQP4 has an association with the immunoregulation of the liver granuloma formation, which may confer a new option for schistosomiasis treatment.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
