

Quantitative analysis of purine nucleotides indicates that purinosomes increase de novo purine biosynthesis
- PMID: 25605736
- PMCID: PMC4358094
- DOI: 10.1074/jbc.M114.628701
Quantitative analysis of purine nucleotides indicates that purinosomes increase de novo purine biosynthesis
Abstract
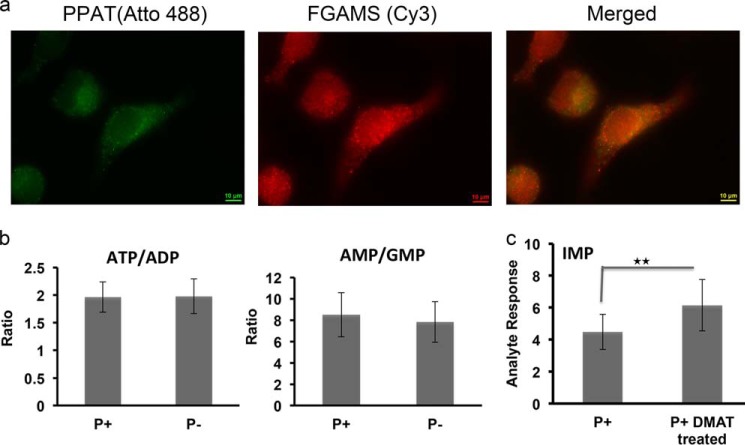
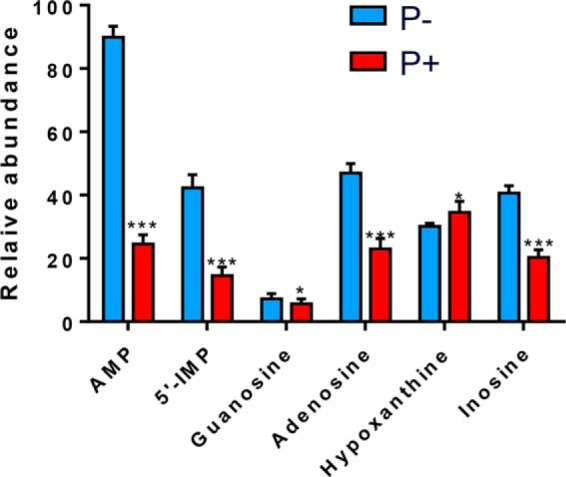
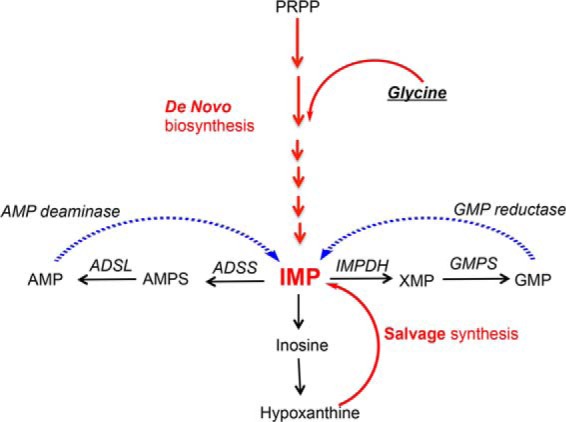
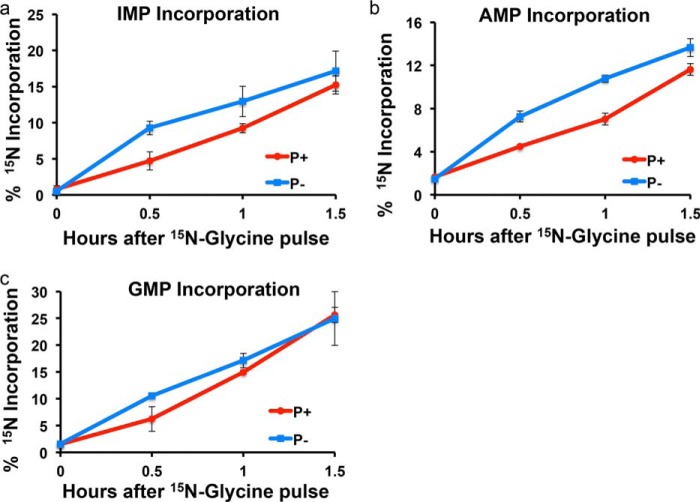
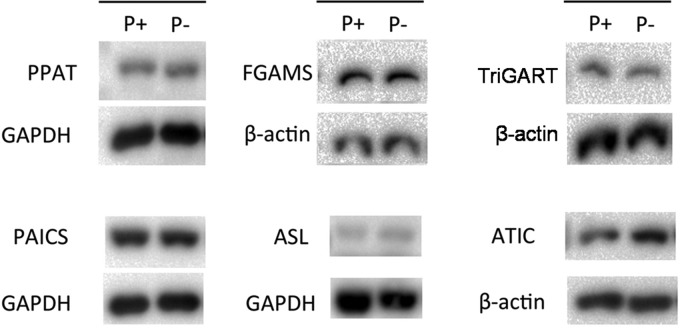
Enzymes in the de novo purine biosynthesis pathway are recruited to form a dynamic metabolic complex referred to as the purinosome. Previous studies have demonstrated that purinosome assembly responds to purine levels in culture medium. Purine-depleted medium or 2-dimethylamino-4,5,6,7-tetrabromo-1H-benzimidazole (DMAT) treatment stimulates the purinosome assembly in HeLa cells. Here, several metabolomic technologies were applied to quantify the static cellular levels of purine nucleotides and measure the de novo biosynthesis rate of IMP, AMP, and GMP. Direct comparison of purinosome-rich cells (cultured in purine-depleted medium) and normal cells showed a 3-fold increase in IMP concentration in purinosome-rich cells and similar levels of AMP, GMP, and ratios of AMP/GMP and ATP/ADP for both. In addition, a higher level of IMP was also observed in HeLa cells treated with DMAT. Furthermore, increases in the de novo IMP/AMP/GMP biosynthetic flux rate under purine-depleted condition were observed. The synthetic enzymes, adenylosuccinate synthase (ADSS) and inosine monophosphate dehydrogenase (IMPDH), downstream of IMP were also shown to be part of the purinosome. Collectively, these results provide further evidence that purinosome assembly is directly related to activated de novo purine biosynthesis, consistent with the functionality of the purinosome.
Keywords: Mass Spectrometry (MS); Metabolic Flux; Metabolomics; Nucleoside/Nucleotide Biosynthesis; Protein Complex; Purine; Purinosome.
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures

References
-
- Rowe P. B., McCairns E., Madsen G., Sauer D., Elliott H. (1978) De novo purine synthesis in avian liver: co-purification of the enzymes and properties of the pathway. J. Biol. Chem. 253, 7711–7721 - PubMed
-
- Rowe P. B., Wyngaarden J. B. (1968) Glutamine phosphoribosylpyrophosphate amidotransferase: purification, substructure, amino acid composition, and absorption spectra. J. Biol. Chem. 243, 6373–6383 - PubMed
-
- Smith G. K., Mueller W. T., Wasserman G. F., Taylor W. D., Benkovic S. J. (1980) Characterization of the enzyme complex involving the folate-requiring enzymes of de novo purine biosynthesis. Biochemistry 19, 4313–4321 - PubMed
-
- Wombacher H. (1983) Molecular compartmentation by enzyme cluster formation: a view over current investigations. Mol. Cell. Biochem. 56, 155–164 - PubMed
-
- Srere P. A. (1987) Complexes of sequential metabolic enzymes. Annu. Rev. Biochem. 56, 89–124 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
