

Alternative splicing regulates the expression of G9A and SUV39H2 methyltransferases, and dramatically changes SUV39H2 functions
- PMID: 25605796
- PMCID: PMC4330376
- DOI: 10.1093/nar/gkv013
Alternative splicing regulates the expression of G9A and SUV39H2 methyltransferases, and dramatically changes SUV39H2 functions
Abstract
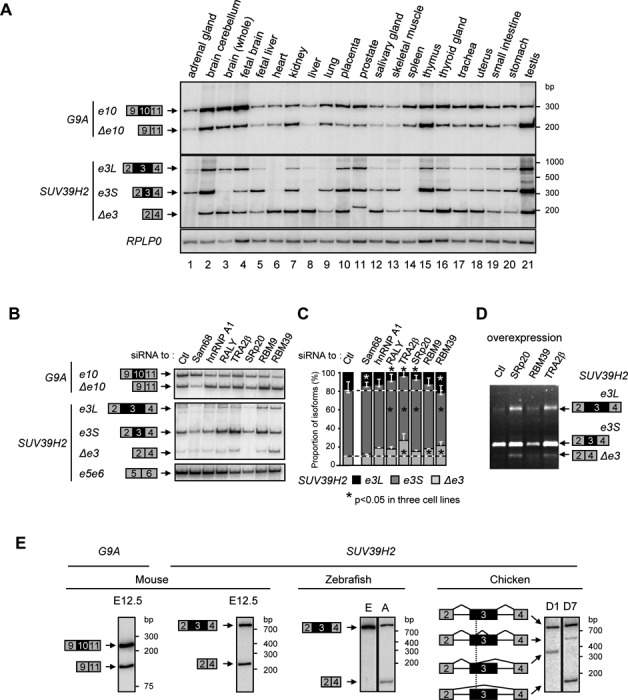
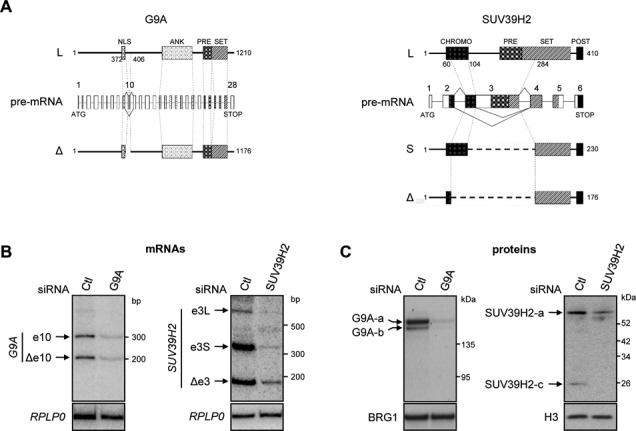
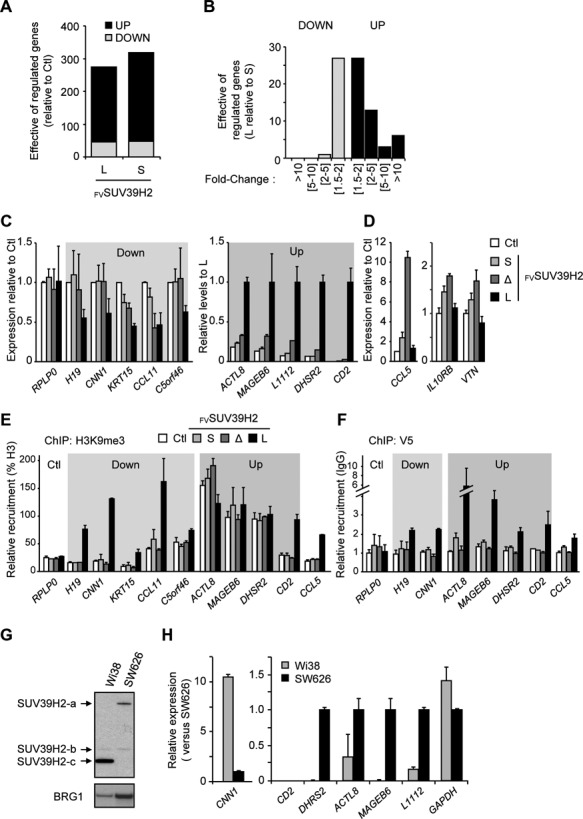
Alternative splicing is the main source of proteome diversity. Here, we have investigated how alternative splicing affects the function of two human histone methyltransferases (HMTase): G9A and SUV39H2. We show that exon 10 in G9A and exon 3 in SUV39H2 are alternatively included in a variety of tissues and cell lines, as well as in a different species. The production of these variants is likely tightly regulated because both constitutive and alternative splicing factors control their splicing profiles. Based on this evidence, we have assessed the link between the inclusion of these exons and the activity of both enzymes. We document that these HMTase genes yield several protein isoforms, which are likely issued from alternative splicing regulation. We demonstrate that inclusion of SUV39H2 exon 3 is a determinant of the stability, the sub-nuclear localization, and the HMTase activity. Genome-wide expression analysis further revealed that alternative inclusion of SUV39H2 exon 3 differentially modulates the expression of target genes. Our data also suggest that a variant of G9A may display a function that is independent of H3K9 methylation. Our work emphasizes that expression and function of genes are not collinear; therefore alternative splicing must be taken into account in any functional study.
© The Author(s) 2015. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures


References
-
- Pan Q., Shai O., Lee L.J., Frey B.J., Blencowe B.J. Deep surveying of alternative splicing complexity in the human transcriptome by high-throughput sequencing. Nat. Genet. 2008;40:1413–1415. - PubMed
-
- Jurica M.S., Moore M.J. Pre-mRNA splicing: awash in a sea of proteins. Mol. Cell. 2003;12:5–14. - PubMed
-
- Wahl M.C., Will C.L., Lührmann R. The spliceosome: design principles of a dynamic RNP machine. Cell. 2009;136:701–718. - PubMed
-
- Krainer A.R., Maniatis T., Ruskin B., Green M.R. Normal and mutant human beta-globin pre-mRNAs are faithfully and efficiently spliced in vitro. Cell. 1984;36:993–1005. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
