

Regulation of glucose tolerance and sympathetic activity by MC4R signaling in the lateral hypothalamus
- PMID: 25605803
- PMCID: PMC4439564
- DOI: 10.2337/db14-1257
Regulation of glucose tolerance and sympathetic activity by MC4R signaling in the lateral hypothalamus
Abstract
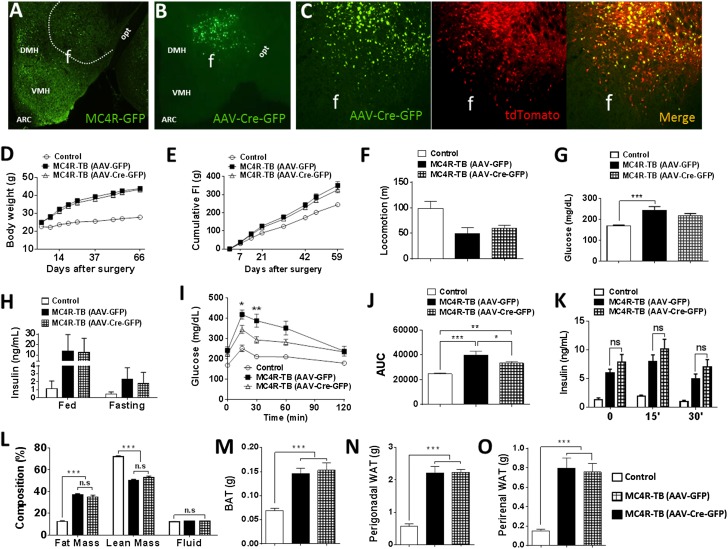
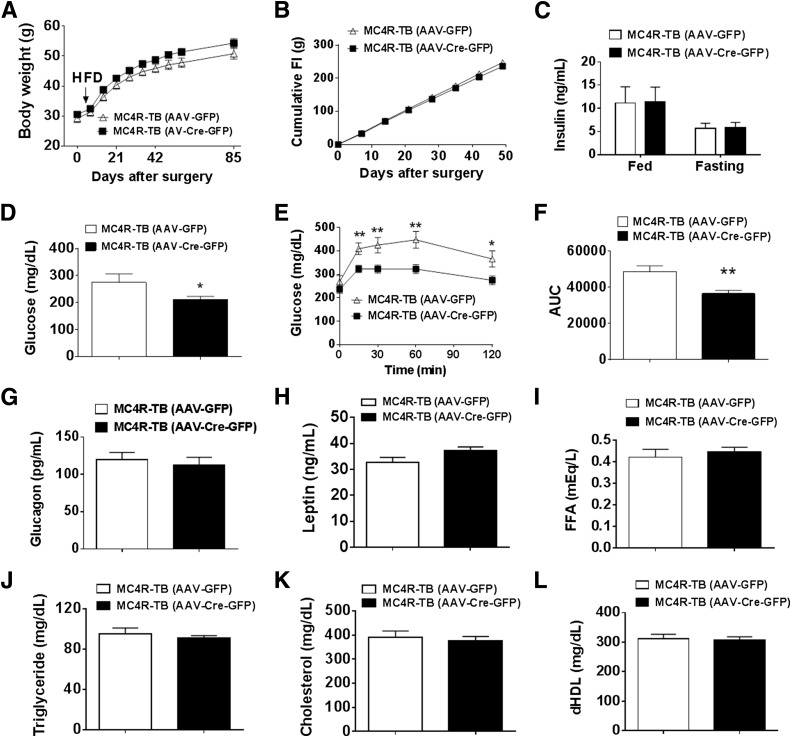
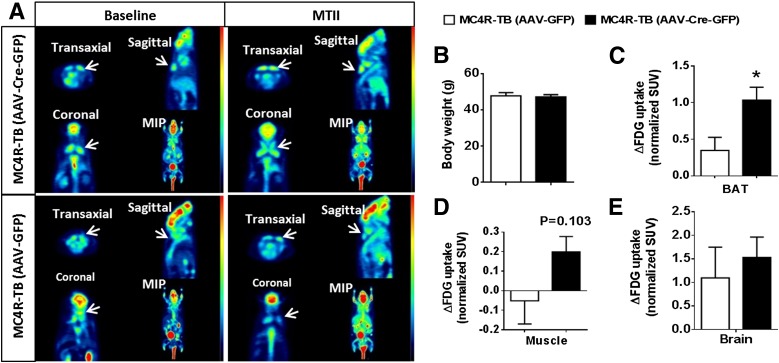
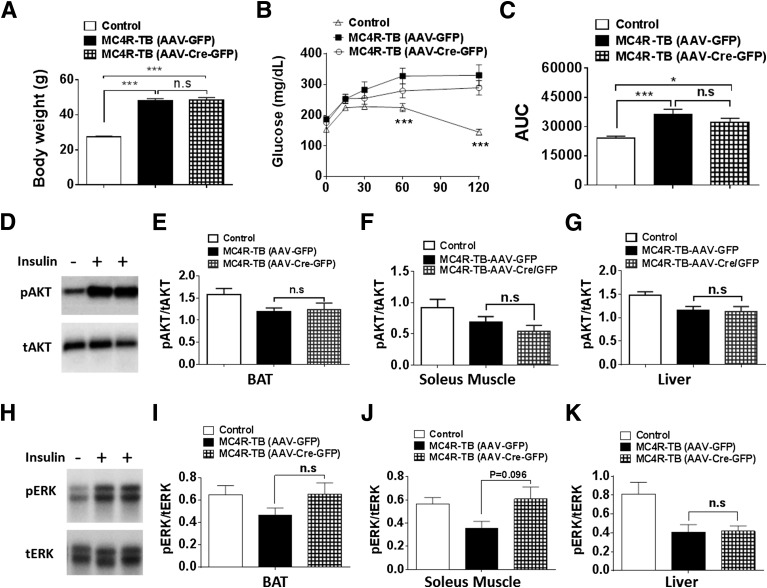
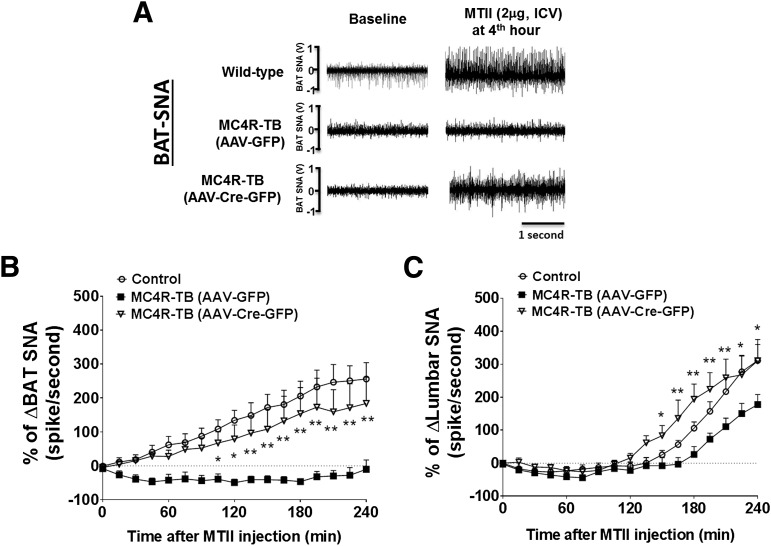
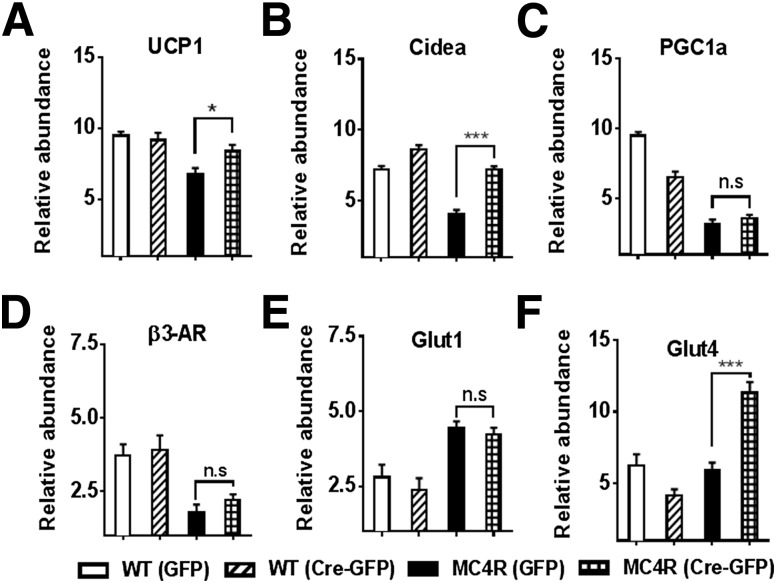
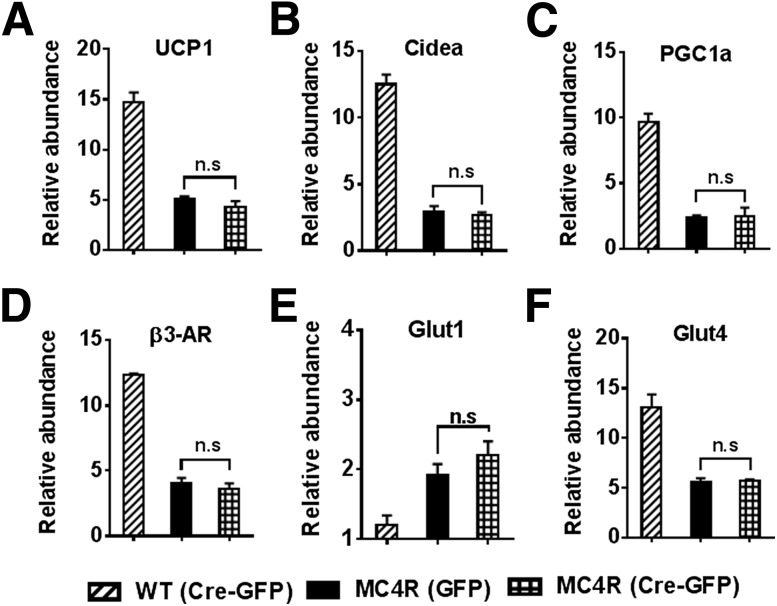
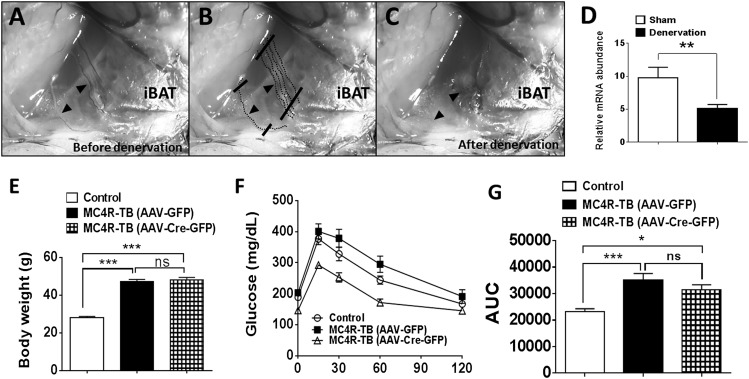
Melanocortin 4 receptor (MC4R) signaling mediates diverse physiological functions, including energy balance, glucose homeostasis, and autonomic activity. Although the lateral hypothalamic area (LHA) is known to express MC4Rs and to receive input from leptin-responsive arcuate proopiomelanocortin neurons, the physiological functions of MC4Rs in the LHA are incompletely understood. We report that MC4R(LHA) signaling regulates glucose tolerance and sympathetic nerve activity. Restoring expression of MC4Rs specifically in the LHA improves glucose intolerance in obese MC4R-null mice without affecting body weight or circulating insulin levels. Fluorodeoxyglucose-mediated tracing of whole-body glucose uptake identifies the interscapular brown adipose tissue (iBAT) as a primary source where glucose uptake is increased in MC4R(LHA) mice. Direct multifiber sympathetic nerve recording further reveals that sympathetic traffic to iBAT is significantly increased in MC4R(LHA) mice, which accompanies a significant elevation of Glut4 expression in iBAT. Finally, bilateral iBAT denervation prevents the glucoregulatory effect of MC4R(LHA) signaling. These results identify a novel role for MC4R(LHA) signaling in the control of sympathetic nerve activity and glucose tolerance independent of energy balance.
© 2015 by the American Diabetes Association. Readers may use this article as long as the work is properly cited, the use is educational and not for profit, and the work is not altered.
Figures
References
-
- Hossain P, Kawar B, El Nahas M. Obesity and diabetes in the developing world—a growing challenge. N Engl J Med 2007;356:213–215 - PubMed
-
- Myers MG Jr, Olson DP. Central nervous system control of metabolism. Nature 2012;491:357–363 - PubMed
-
- Kleinridders A, Könner AC, Brüning JC. CNS-targets in control of energy and glucose homeostasis. Curr Opin Pharmacol 2009;9:794–804 - PubMed
-
- Cone RD. Anatomy and regulation of the central melanocortin system. Nat Neurosci 2005;8:571–578 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
