

Biocontainment of genetically modified organisms by synthetic protein design
- PMID: 25607366
- PMCID: PMC4422498
- DOI: 10.1038/nature14121
Biocontainment of genetically modified organisms by synthetic protein design
Erratum in
-
Corrigendum: Biocontainment of genetically modified organisms by synthetic protein design.Nature. 2015 Nov 12;527(7577):264. doi: 10.1038/nature15536. Epub 2015 Sep 23. Nature. 2015. PMID: 26416745 No abstract available.
Abstract
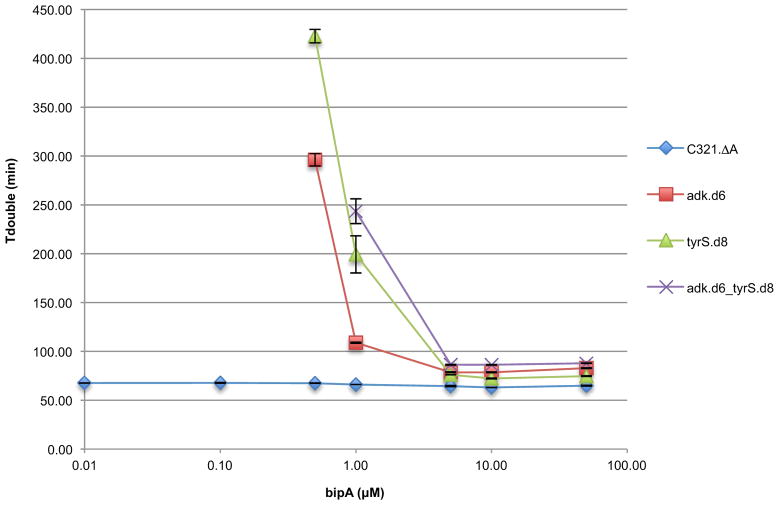
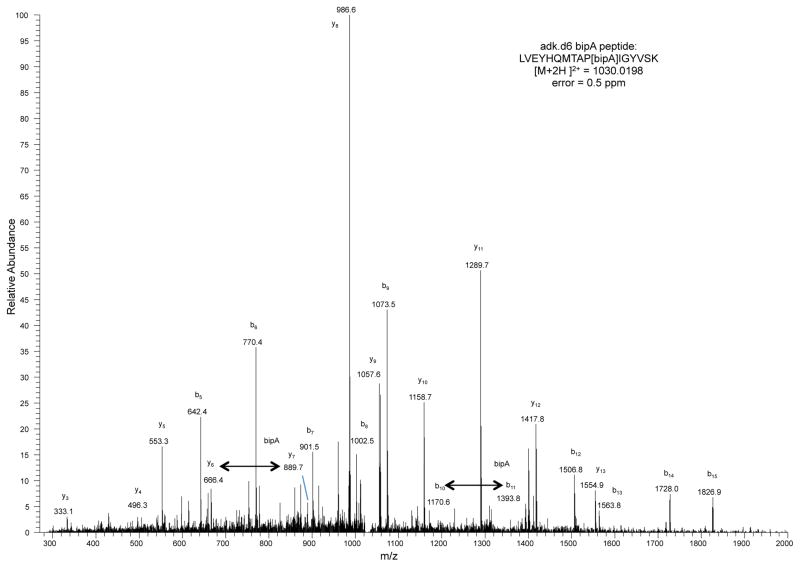
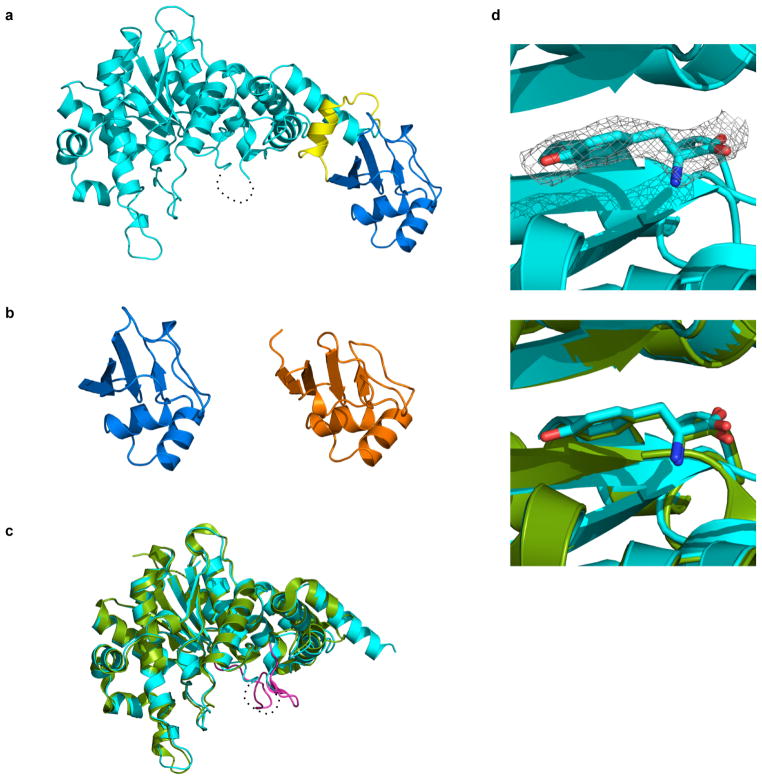
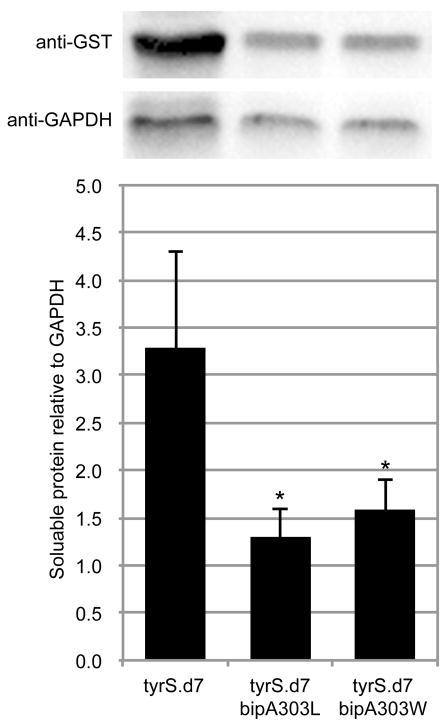
Genetically modified organisms (GMOs) are increasingly deployed at large scales and in open environments. Genetic biocontainment strategies are needed to prevent unintended proliferation of GMOs in natural ecosystems. Existing biocontainment methods are insufficient because they impose evolutionary pressure on the organism to eject the safeguard by spontaneous mutagenesis or horizontal gene transfer, or because they can be circumvented by environmentally available compounds. Here we computationally redesign essential enzymes in the first organism possessing an altered genetic code (Escherichia coli strain C321.ΔA) to confer metabolic dependence on non-standard amino acids for survival. The resulting GMOs cannot metabolically bypass their biocontainment mechanisms using known environmental compounds, and they exhibit unprecedented resistance to evolutionary escape through mutagenesis and horizontal gene transfer. This work provides a foundation for safer GMOs that are isolated from natural ecosystems by a reliance on synthetic metabolites.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
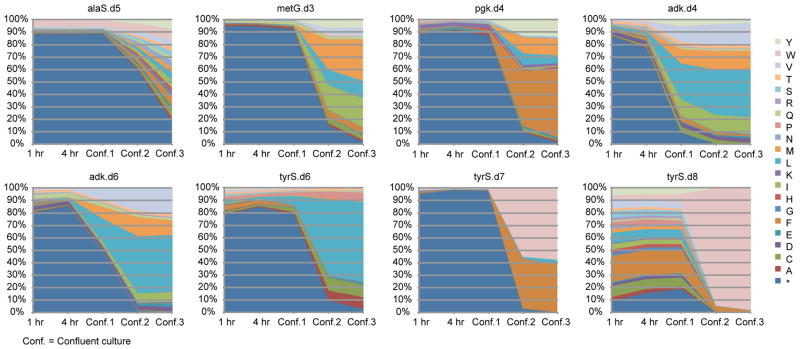
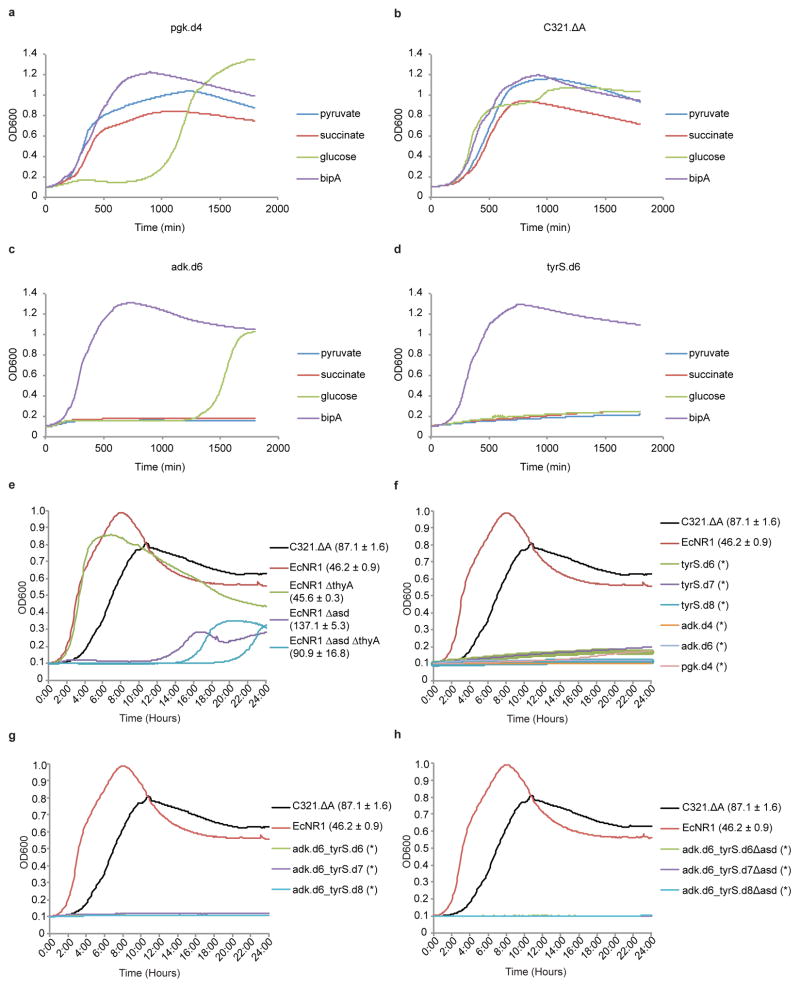
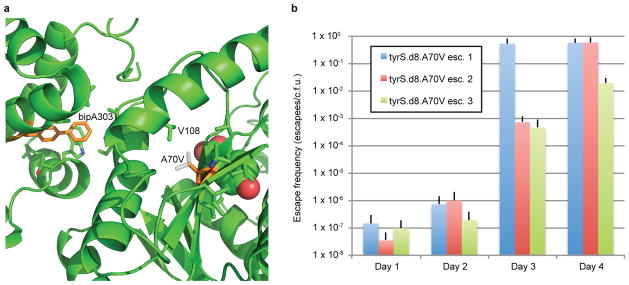
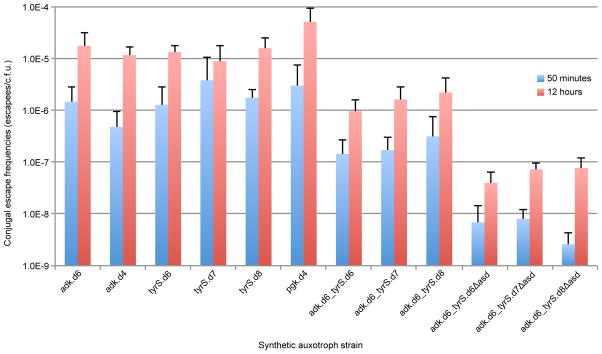
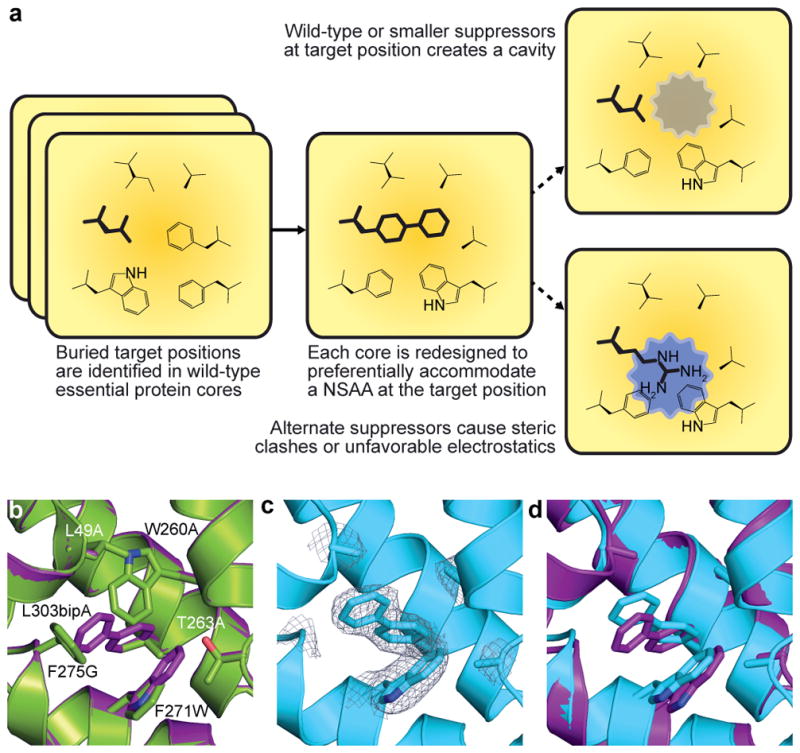
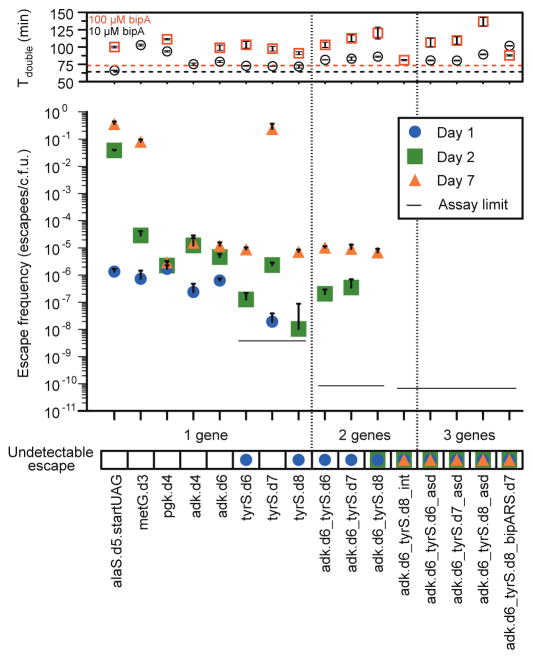
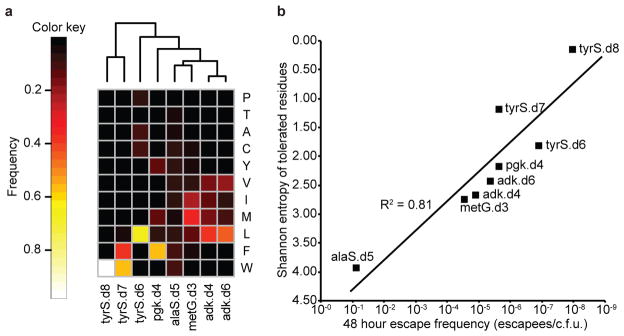
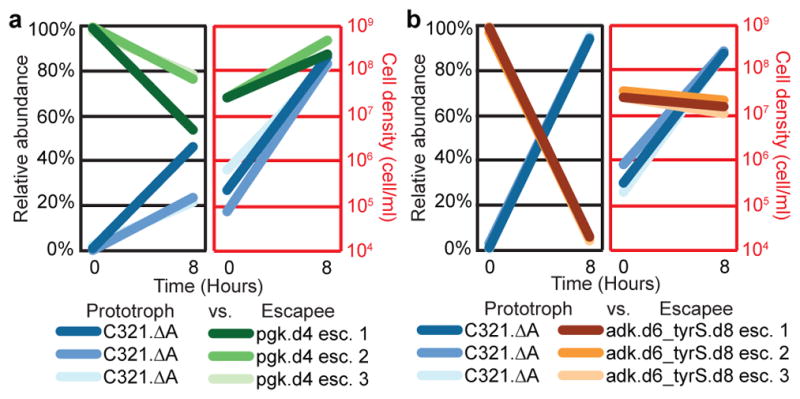
Comment in
-
Synthetic biology: GMOs in lockdown.Nat Rev Microbiol. 2015 Mar;13(3):125. doi: 10.1038/nrmicro3443. Epub 2015 Feb 9. Nat Rev Microbiol. 2015. PMID: 25659321 No abstract available.
-
Synthetic biology: GMOs in lockdown.Nat Rev Genet. 2015 Mar;16(3):127. doi: 10.1038/nrg3909. Nat Rev Genet. 2015. PMID: 25690387 No abstract available.
References
-
- Molin S, et al. Conditional Suicide System for Containment of Bacteria and Plasmids. Nat Biotech. 1987;5:1315–1318.
-
- Curtiss R., 3rd Biological containment and cloning vector transmissibility. The Journal of infectious diseases. 1978;137:668–675. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
