

Characterization of hepatitis C virus interaction with heparan sulfate proteoglycans
- PMID: 25609801
- PMCID: PMC4403428
- DOI: 10.1128/JVI.03647-14
Characterization of hepatitis C virus interaction with heparan sulfate proteoglycans
Abstract
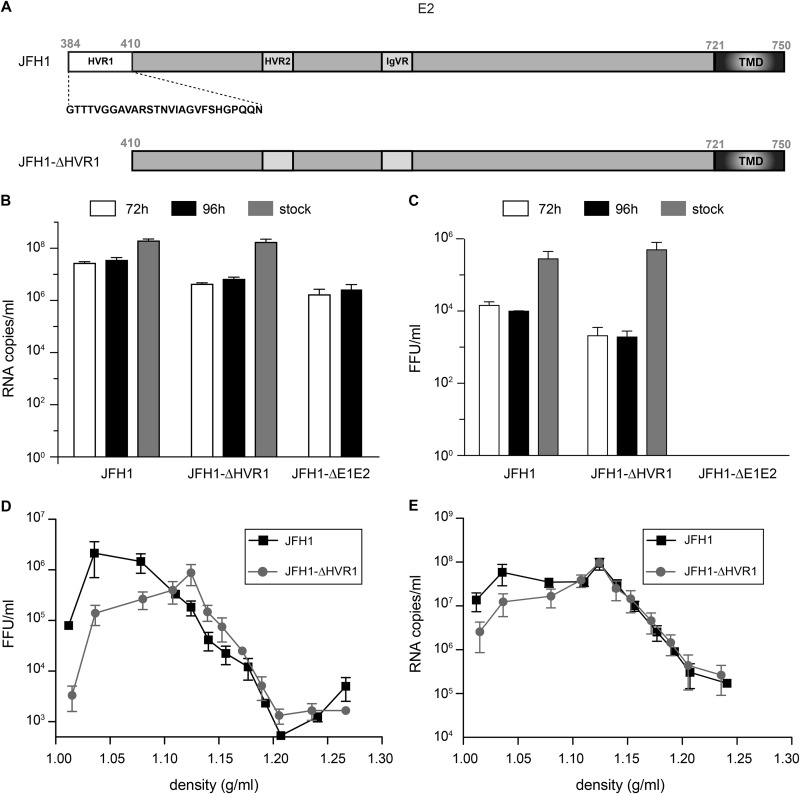
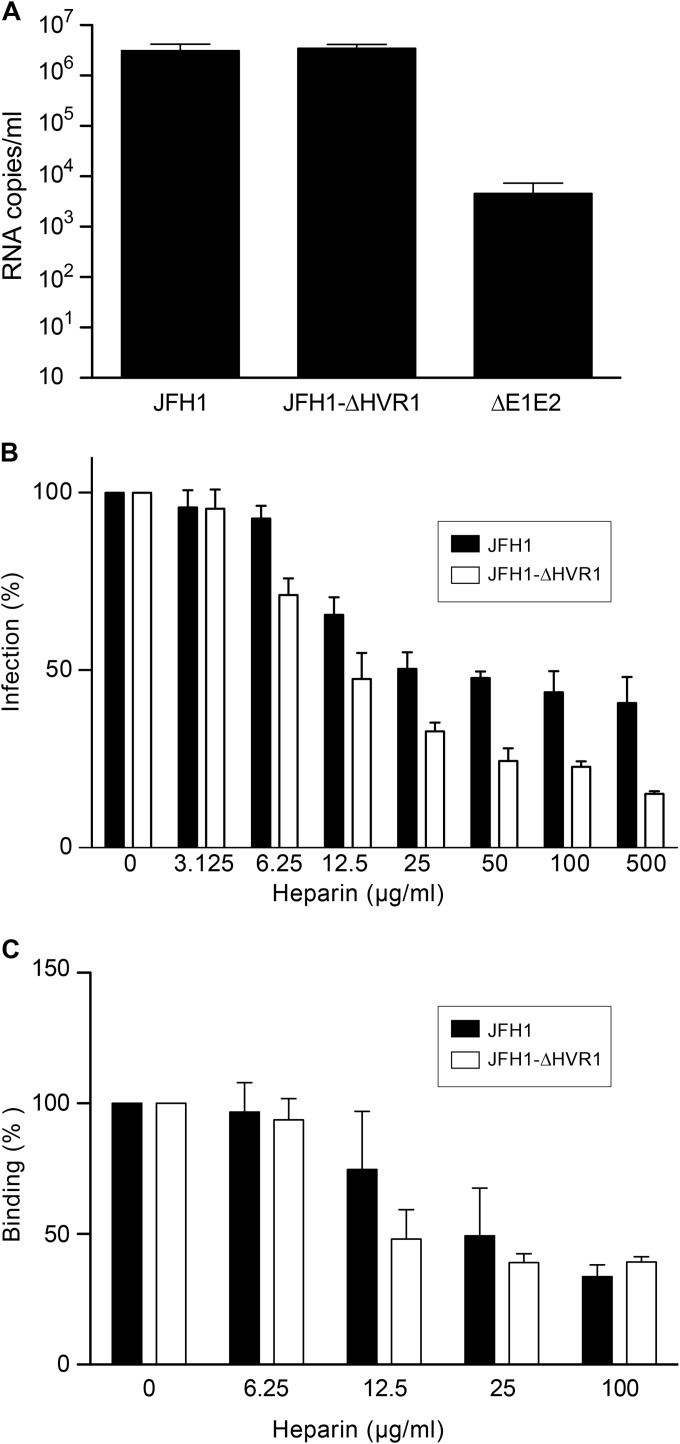
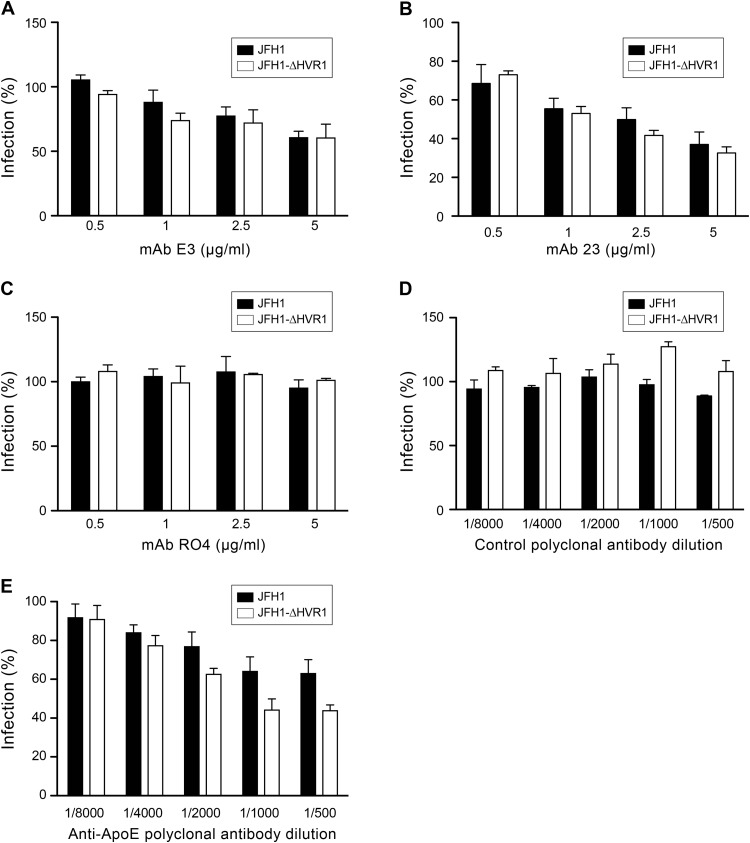
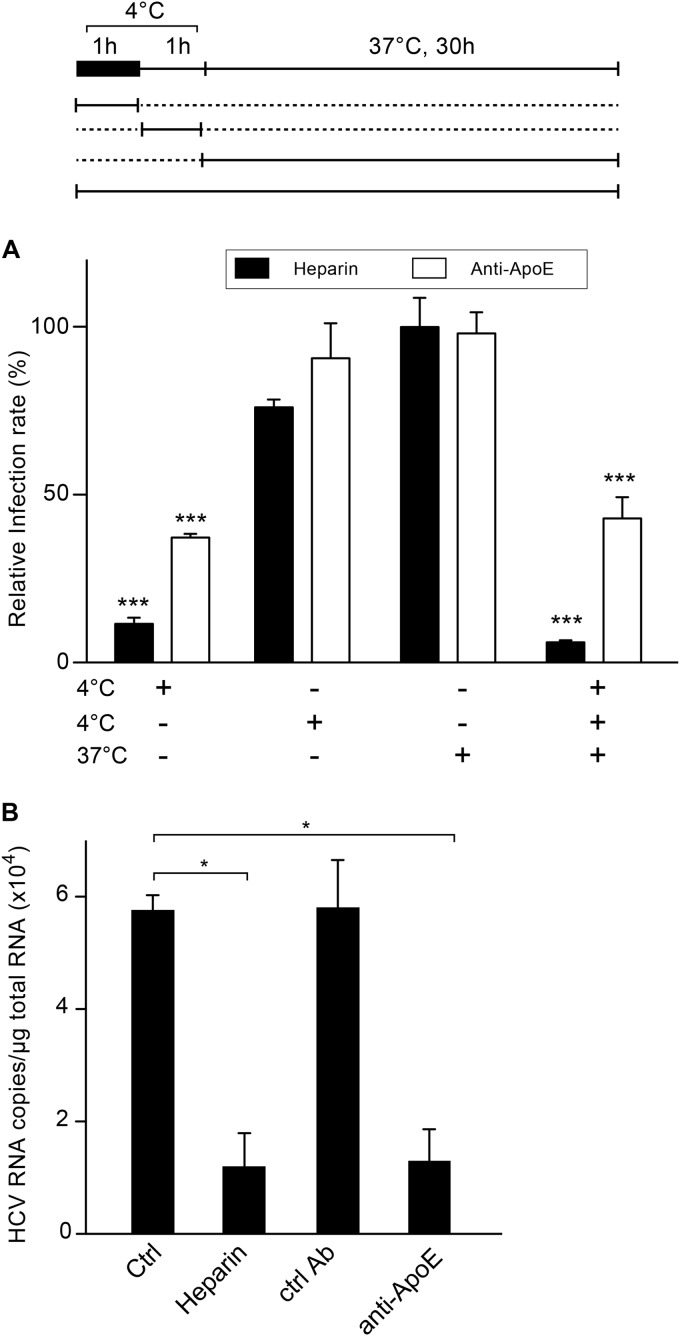
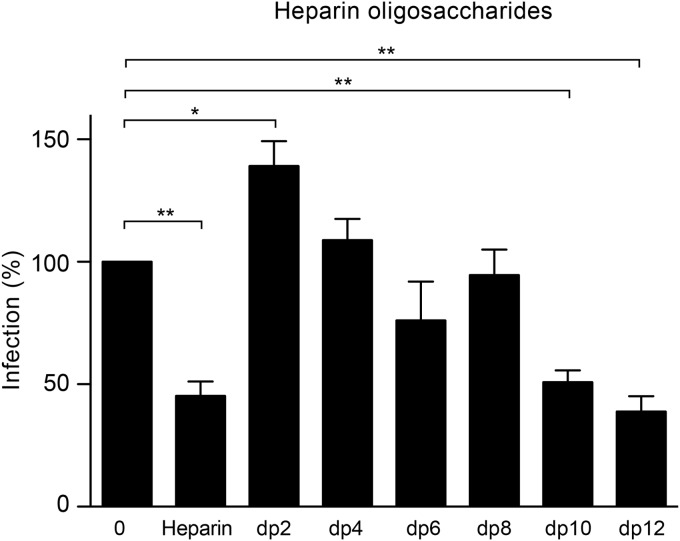
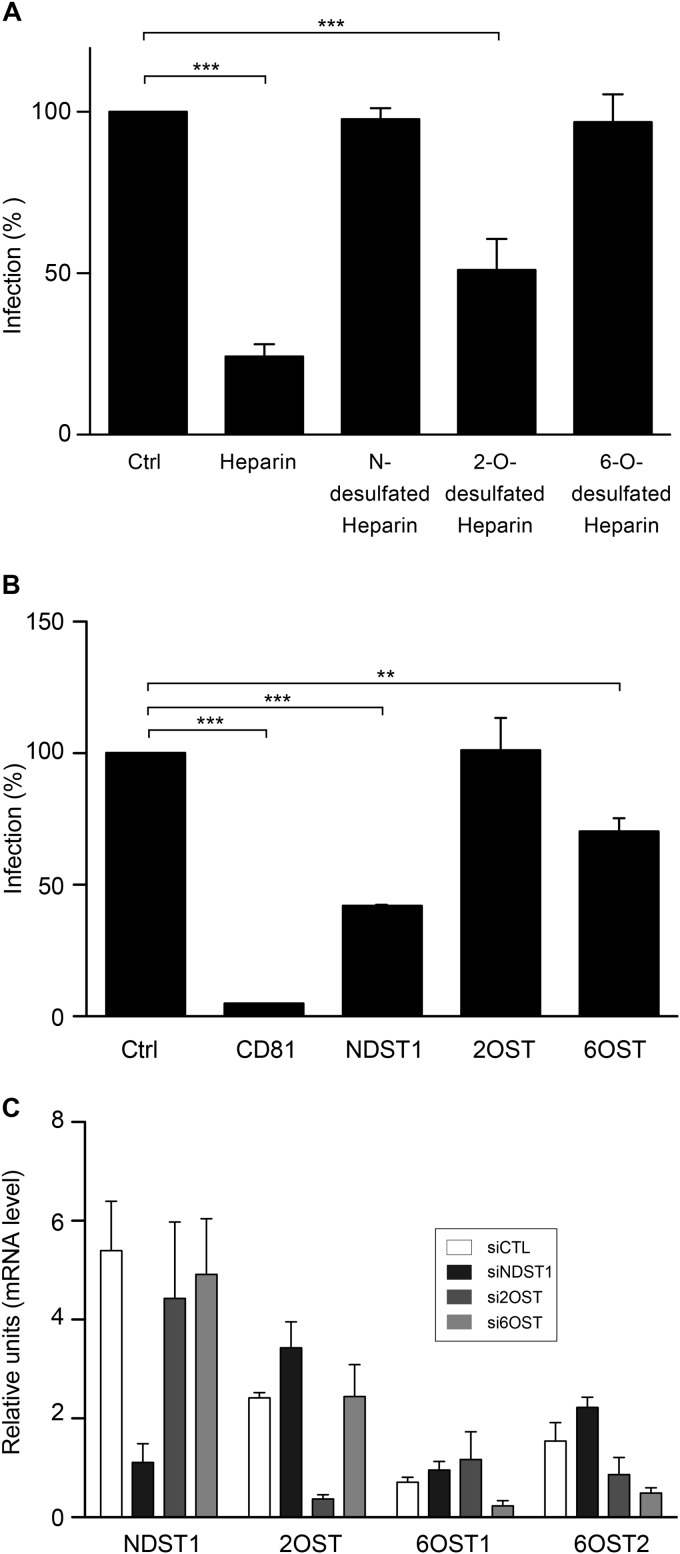
Hepatitis C virus (HCV) entry involves binding to cell surface heparan sulfate (HS) structures. However, due to the lipoprotein-like structure of HCV, the exact contribution of virion components to this interaction remains controversial. Here, we investigated the relative contribution of HCV envelope proteins and apolipoprotein E in the HS-binding step. Deletion of hypervariable region 1, a region previously proposed to be involved in HS binding, did not alter HCV virion binding to HS, indicating that this region is not involved in this interaction in the context of a viral infection. Patient sera and monoclonal antibodies recognizing different regions of HCV envelope glycoproteins were also used in a pulldown assay with beads coated with heparin, a close HS structural homologue. Although isolated HCV envelope glycoproteins could interact with heparin, none of these antibodies was able to interfere with the virion-heparin interaction, strongly suggesting that at the virion surface, HCV envelope glycoproteins are not accessible for HS binding. In contrast, results from kinetic studies, heparin pulldown experiments, and inhibition experiments with anti-apolipoprotein E antibodies indicated that this apolipoprotein plays a major role in HCV-HS interaction. Finally, characterization of the HS structural determinants required for HCV infection by silencing of the enzymes involved in the HS biosynthesis pathway and by competition with modified heparin indicated that N- and 6-O-sulfation but not 2-O-sulfation is required for HCV infection and that the minimum HS oligosaccharide length required for HCV infection is a decasaccharide. Together, these data indicate that HCV hijacks apolipoprotein E to initiate its interaction with specific HS structures.
Importance: Hepatitis C is a global health problem. Hepatitis C virus (HCV) infects approximately 130 million individuals worldwide, with the majority of cases remaining undiagnosed and untreated. In most infected individuals, the virus evades the immune system and establishes a chronic infection. As a consequence, hepatitis C is the leading cause of cirrhosis, end-stage liver disease, hepatocellular carcinoma, and liver transplantation. Virus infection is initiated by entry of the virus into the host cell. In this study, we provide new insights into the viral and cellular determinants involved in the first step of HCV entry, the binding of the virus to host cells. We show that apolipoprotein E is likely responsible for virus binding to heparan sulfate and that N- and 6-O-sulfation of the heparan sulfate proteoglycans is required for HCV infection. In addition, the minimal HS length unit required for HCV infection is a decasaccharide.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures



References
-
- Ray SC, Bailey JR, Thomas DL. 2013. Hepatitis C virus. InKnipe DM, Howley PM, Cohen JI, Griffin DE, Lamb RA, Martin MA, Racaniello VR, Roizman B (ed), Fields virology, 6th ed (electronic) Lippincott Wilkins & Williams, Philadelphia, PA.
-
- Bartosch B, Vitelli A, Granier C, Goujon C, Dubuisson J, Pascale S, Scarselli E, Cortese R, Nicosia A, Cosset FL. 2003. Cell entry of hepatitis C virus requires a set of co-receptors that include the CD81 tetraspanin and the SR-B1 scavenger receptor. J Biol Chem 278:41624–41630. doi: 10.1074/jbc.M305289200. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
