

Polarized cellular patterns of endocannabinoid production and detection shape cannabinoid signaling in neurons
- PMID: 25610369
- PMCID: PMC4285097
- DOI: 10.3389/fncel.2014.00426
Polarized cellular patterns of endocannabinoid production and detection shape cannabinoid signaling in neurons
Abstract
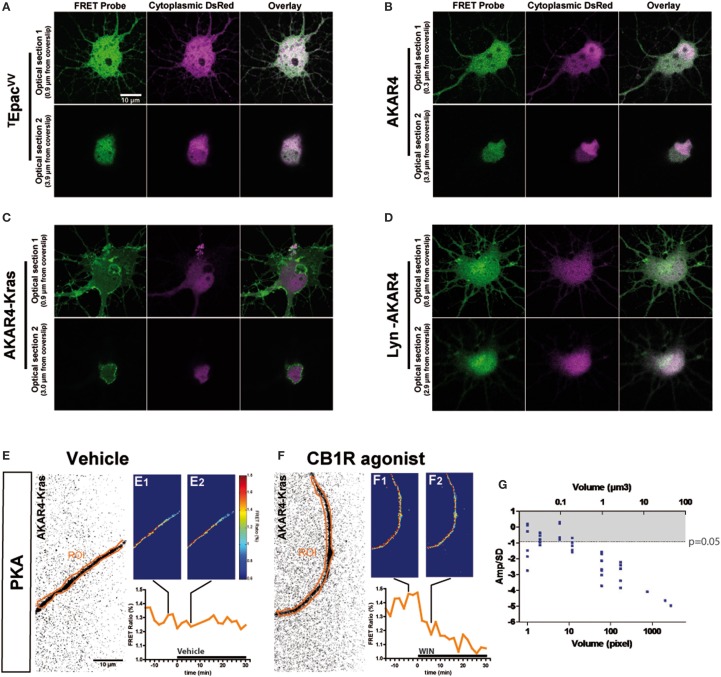
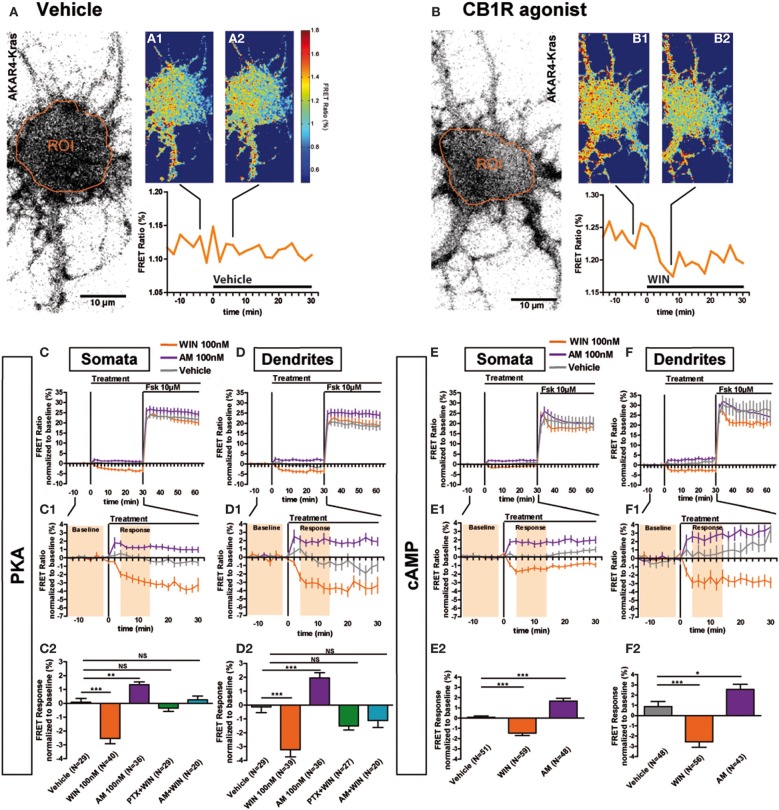
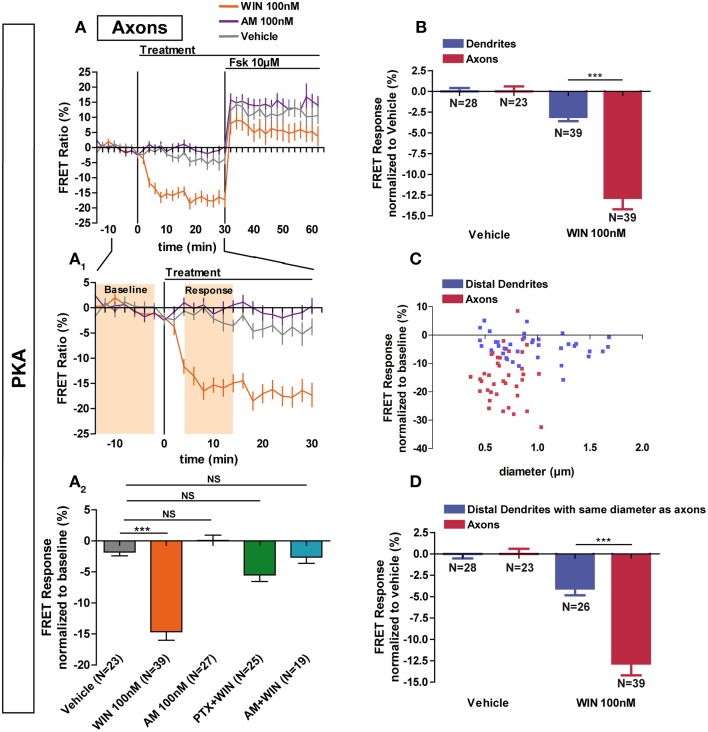
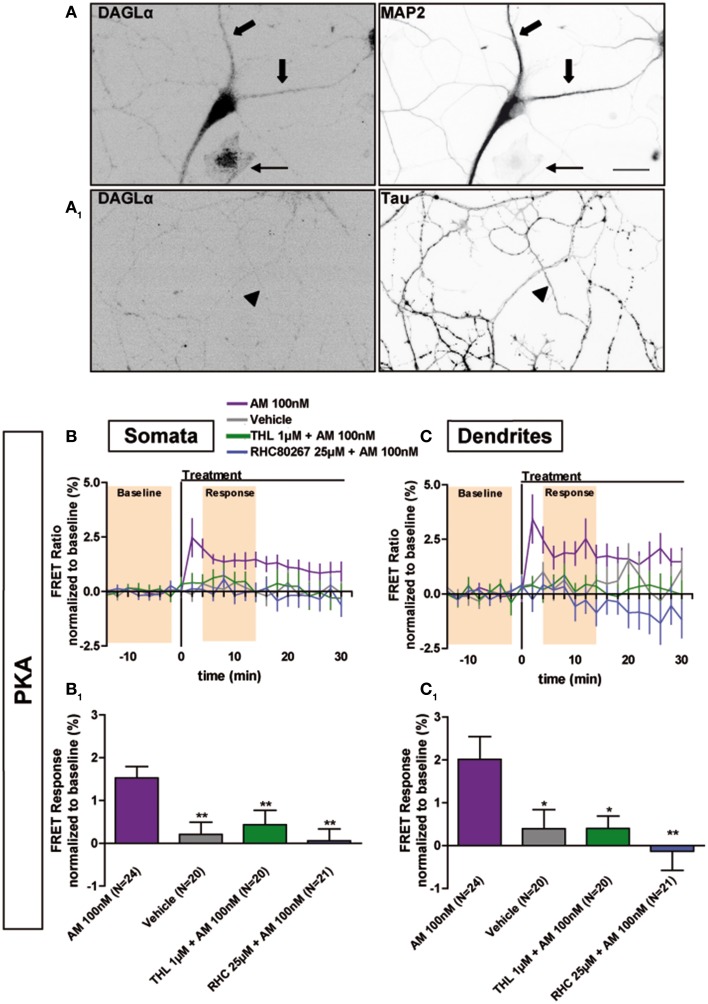
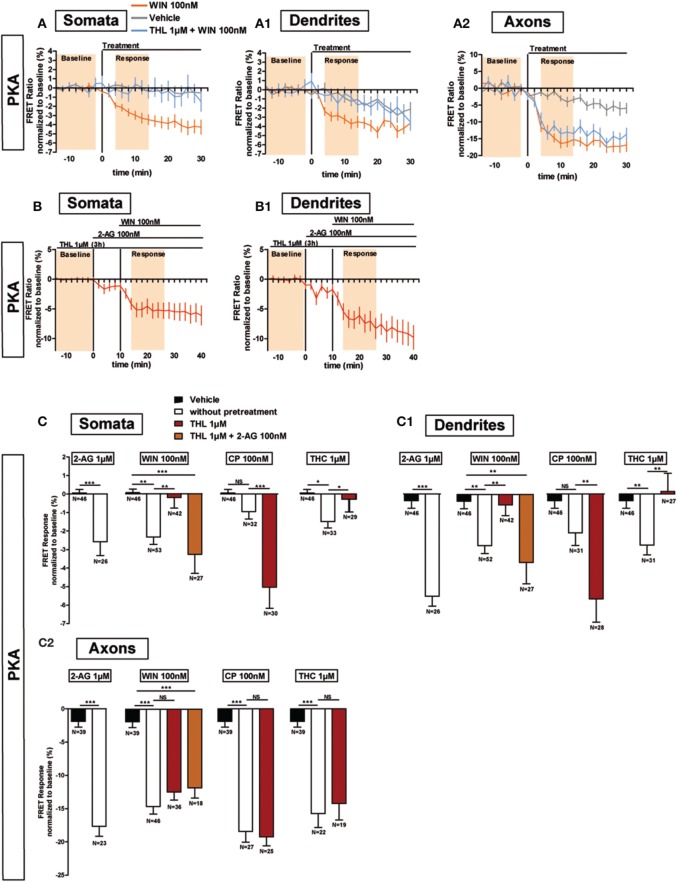
Neurons display important differences in plasma membrane composition between somatodendritic and axonal compartments, potentially leading to currently unexplored consequences in G-protein-coupled-receptor signaling. Here, by using highly-resolved biosensor imaging to measure local changes in basal levels of key signaling components, we explored features of type-1 cannabinoid receptor (CB1R) signaling in individual axons and dendrites of cultured rat hippocampal neurons. Activation of endogenous CB1Rs led to rapid, Gi/o-protein- and cAMP-mediated decrease of cyclic-AMP-dependent protein kinase (PKA) activity in the somatodendritic compartment. In axons, PKA inhibition was significantly stronger, in line with axonally-polarized distribution of CB1Rs. Conversely, inverse agonist AM281 produced marked rapid increase of basal PKA activation in somata and dendrites, but not in axons, removing constitutive activation of CB1Rs generated by local production of the endocannabinoid 2-arachidonoylglycerol (2-AG). Interestingly, somatodendritic 2-AG levels differently modified signaling responses to CB1R activation by Δ(9)-THC, the psychoactive compound of marijuana, and by the synthetic cannabinoids WIN55,212-2 and CP55,940. These highly contrasted differences in sub-neuronal signaling responses warrant caution in extrapolating pharmacological profiles, which are typically obtained in non-polarized cells, to predict in vivo responses of axonal (i.e., presynaptic) GPCRs. Therefore, our results suggest that enhanced comprehension of GPCR signaling constraints imposed by neuronal cell biology may improve the understanding of neuropharmacological action.
Keywords: CB1; DAGL; FRET; allosteric; biased agonism; cyclic nucleotide; endocannabinoid; lipid.
Figures
Similar articles
-
Constitutive activation drives compartment-selective endocytosis and axonal targeting of type 1 cannabinoid receptors.J Neurosci. 2006 Mar 22;26(12):3141-53. doi: 10.1523/JNEUROSCI.5437-05.2006. J Neurosci. 2006. PMID: 16554465 Free PMC article.
-
Activation-dependent plasticity of polarized GPCR distribution on the neuronal surface.J Mol Cell Biol. 2013 Aug;5(4):250-65. doi: 10.1093/jmcb/mjt014. Epub 2013 Apr 11. J Mol Cell Biol. 2013. PMID: 23585691
-
Activation-dependent subcellular distribution patterns of CB1 cannabinoid receptors in the rat forebrain.Cereb Cortex. 2013 Nov;23(11):2581-91. doi: 10.1093/cercor/bhs240. Epub 2012 Aug 14. Cereb Cortex. 2013. PMID: 22892424
-
Type-1 cannabinoid receptor signaling in neuronal development.Pharmacology. 2012;90(1-2):19-39. doi: 10.1159/000339075. Epub 2012 Jul 3. Pharmacology. 2012. PMID: 22776780 Review.
-
Cannabinoid receptor signaling in progenitor/stem cell proliferation and differentiation.Prog Lipid Res. 2013 Oct;52(4):633-50. doi: 10.1016/j.plipres.2013.05.004. Epub 2013 Sep 25. Prog Lipid Res. 2013. PMID: 24076098 Review.
Cited by
-
Inhibition of 2-AG hydrolysis alleviates posttraumatic headache attributed to mild traumatic brain injury.J Headache Pain. 2024 Jul 16;25(1):115. doi: 10.1186/s10194-024-01817-z. J Headache Pain. 2024. PMID: 39014318 Free PMC article.
-
Protein Interactors and Trafficking Pathways That Regulate the Cannabinoid Type 1 Receptor (CB1R).Front Mol Neurosci. 2020 Jun 12;13:108. doi: 10.3389/fnmol.2020.00108. eCollection 2020. Front Mol Neurosci. 2020. PMID: 32595453 Free PMC article. Review.
-
Editorial: Dynamics of cyclic nucleotide signaling in neurons.Front Cell Neurosci. 2015 Aug 3;9:296. doi: 10.3389/fncel.2015.00296. eCollection 2015. Front Cell Neurosci. 2015. PMID: 26283926 Free PMC article. No abstract available.
-
The Cannabinoid Receptor CB1 Interacts with the WAVE1 Complex and Plays a Role in Actin Dynamics and Structural Plasticity in Neurons.PLoS Biol. 2015 Oct 23;13(10):e1002286. doi: 10.1371/journal.pbio.1002286. eCollection 2015 Oct. PLoS Biol. 2015. PMID: 26496209 Free PMC article.
-
The C-terminal helix 9 motif in rat cannabinoid receptor type 1 regulates axonal trafficking and surface expression.Elife. 2019 Apr 30;8:e44252. doi: 10.7554/eLife.44252. Elife. 2019. PMID: 31036155 Free PMC article.
References
-
- Castro L. R., Gervasi N., Guiot E., Cavellini L., Nikolaev V. O., Paupardin-Tritsch D., et al. . (2010). Type 4 phosphodiesterase plays different integrating roles in different cellular domains in pyramidal cortical neurons. J. Neurosci. 30, 6143–6151. 10.1523/JNEUROSCI.5851-09.2010 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources