

GATA-Binding Factor 6 Contributes to Atrioventricular Node Development and Function
- PMID: 25613430
- PMCID: PMC4406809
- DOI: 10.1161/CIRCGENETICS.113.000587
GATA-Binding Factor 6 Contributes to Atrioventricular Node Development and Function
Abstract
Background: Several transcription factors regulate cardiac conduction system (CCS) development and function but the role of each in specifying distinct CCS components remains unclear. GATA-binding factor 6 (GATA6) is a zinc-finger transcription factor that is critical for patterning the cardiovascular system. However, the role of GATA6 in the embryonic heart and CCS has never been shown.
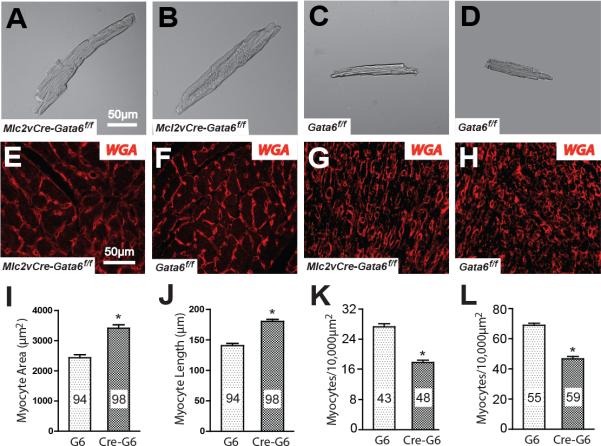
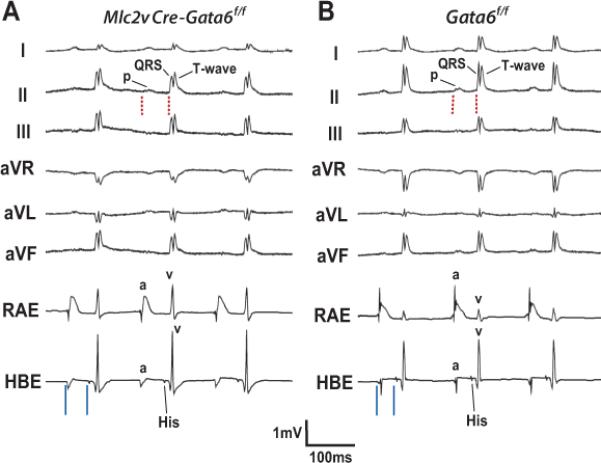
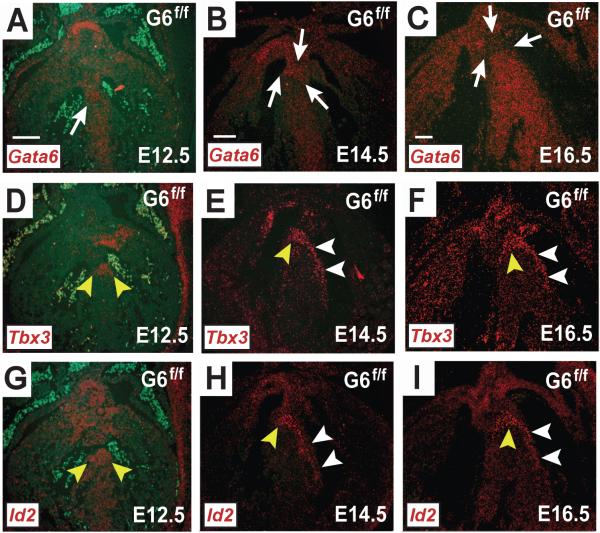
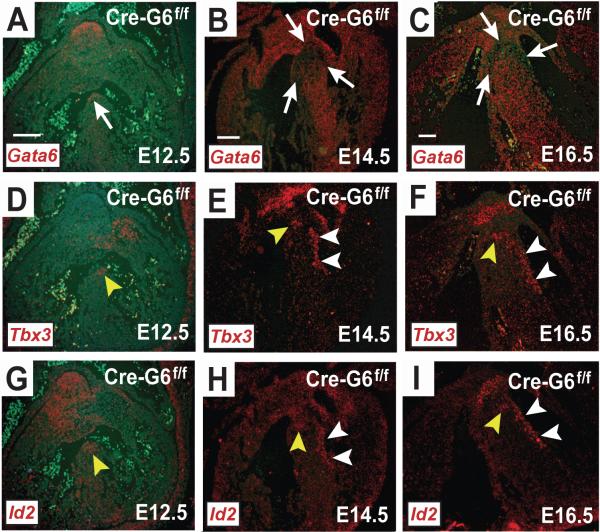
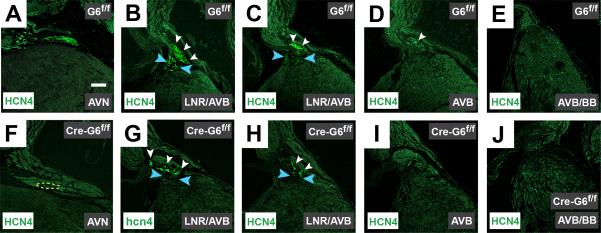
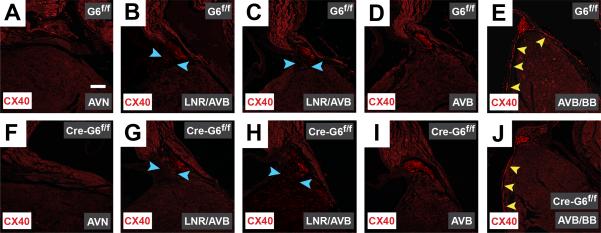
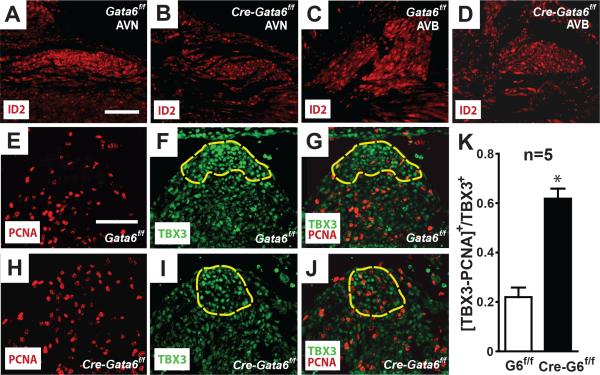
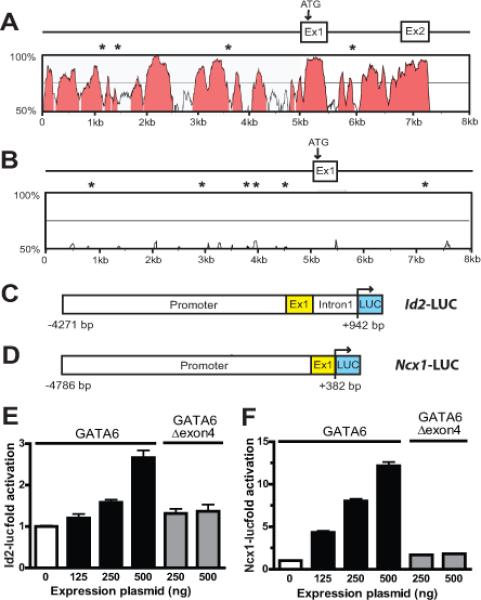
Methods and results: We report that Gata6 is expressed abundantly in the proximal CCS during midgestation in mice. Myocardial-specific deletion of the carboxyl zinc-finger of Gata6 induces loss of hyperpolarizing cyclic nucleotide-gated channel, subtype 4 staining in the compact atrioventricular node with some retention of hyperpolarizing cyclic nucleotide-gated channel, subtype 4 staining in the atrioventricular bundle, but has no significant effect on the connexin-40-positive bundle branches. Furthermore, myocardial-specific deletion of the carboxyl zinc-finger of Gata6 alters atrioventricular conduction in postnatal life as assessed by surface and invasive electrophysiological evaluation, as well as decreasing the number of ventricular myocytes and inducing compensatory myocyte hypertrophy. Myocardial-specific deletion of the carboxyl zinc-finger of Gata6 is also associated with downregulation of the transcriptional repressor ID2 and the cardiac sodium-calcium exchanger NCX1 in the proximal CCS, where GATA6 transactivates both of these factors. Finally, carboxyl zinc-finger deletion of Gata6 reduces cell-cycle exit of TBX3+ myocytes in the developing atrioventricular bundle during the period of atrioventricular node specification, which results in fewer TBX3+ cells in the proximal CCS of mature mutant mice.
Conclusions: GATA6 contributes to the development and postnatal function of the murine atrioventricular node by promoting cell-cycle exit of specified cardiomyocytes toward a conduction system lineage.
Keywords: GATA6 transcription factor; atrioventricular node; cardiac electrophysiology.
© 2015 American Heart Association, Inc.
Figures
Comment in
-
Guiding Cardiac Conduction With GATA.Circ Cardiovasc Genet. 2015 Apr;8(2):247-9. doi: 10.1161/CIRCGENETICS.115.001039. Circ Cardiovasc Genet. 2015. PMID: 25901035 No abstract available.
References
-
- Pashmforoush M, Lu JT, Chen H, Amand TS, Kondo R, Pradervand S, et al. Nkx2-5 pathways and congenital heart disease: loss of ventricular myocyte lineage specification leads to progressive cardiomyopathy and complete heart block. Cell. 2004;117:378–386. - PubMed
-
- Moskowitz IP, Kim JB, Moore ML, Wolf CM, Peterson MA, Shendure J, et al. A molecular pathway including Id2, Tbx5, and Nkx2-5 required for cardiac conduction system development. Cell. 2007;129:1365–1376. - PubMed
-
- Bakker ML, Boukens BJ, Mommersteeg MT, Brons JF, Wakker V, Moorman AF, et al. Transcription factor Tbx3 is required for the specification of the atrioventricular conduction system. Circ Res. 2008;102:1340–1349. - PubMed
-
- Aanhaanen WT, Brons JF, Domínguez JN, Rana MS, Norden J, Airik R, et al. The Tbx2+ primary myocardium of the atrioventricular canal forms the atrioventricular node and the base of the left ventricle. Circ Res. 2009;104:1264–1274. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
