

microRNAs regulate β-catenin of the Wnt signaling pathway in early sea urchin development
- PMID: 25614238
- PMCID: PMC4433612
- DOI: 10.1016/j.ydbio.2015.01.008
microRNAs regulate β-catenin of the Wnt signaling pathway in early sea urchin development
Abstract
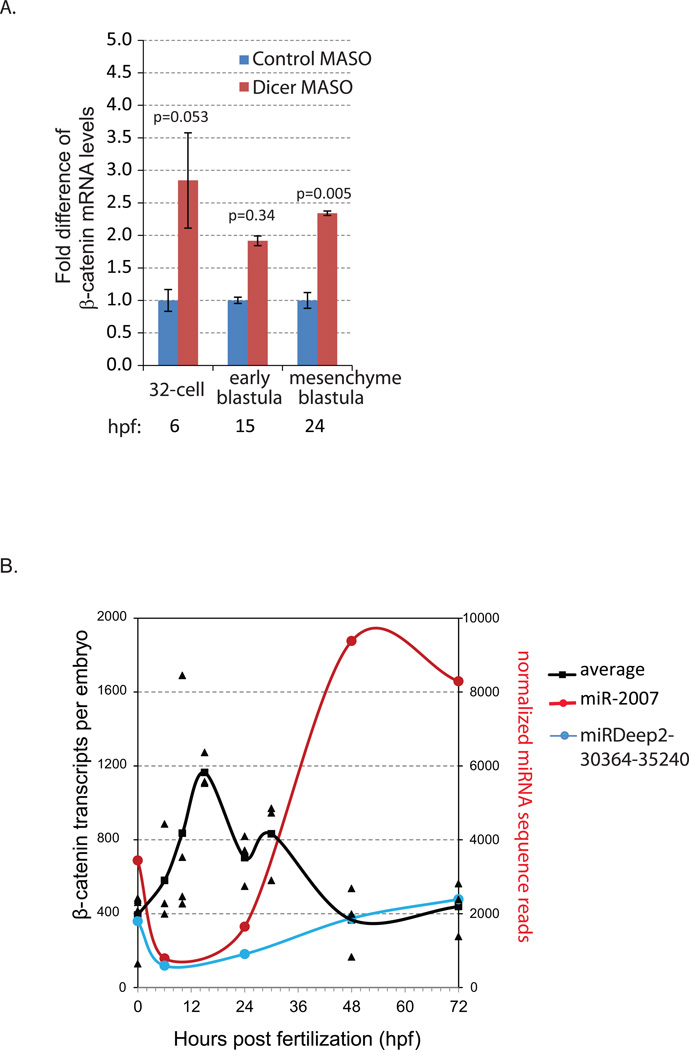
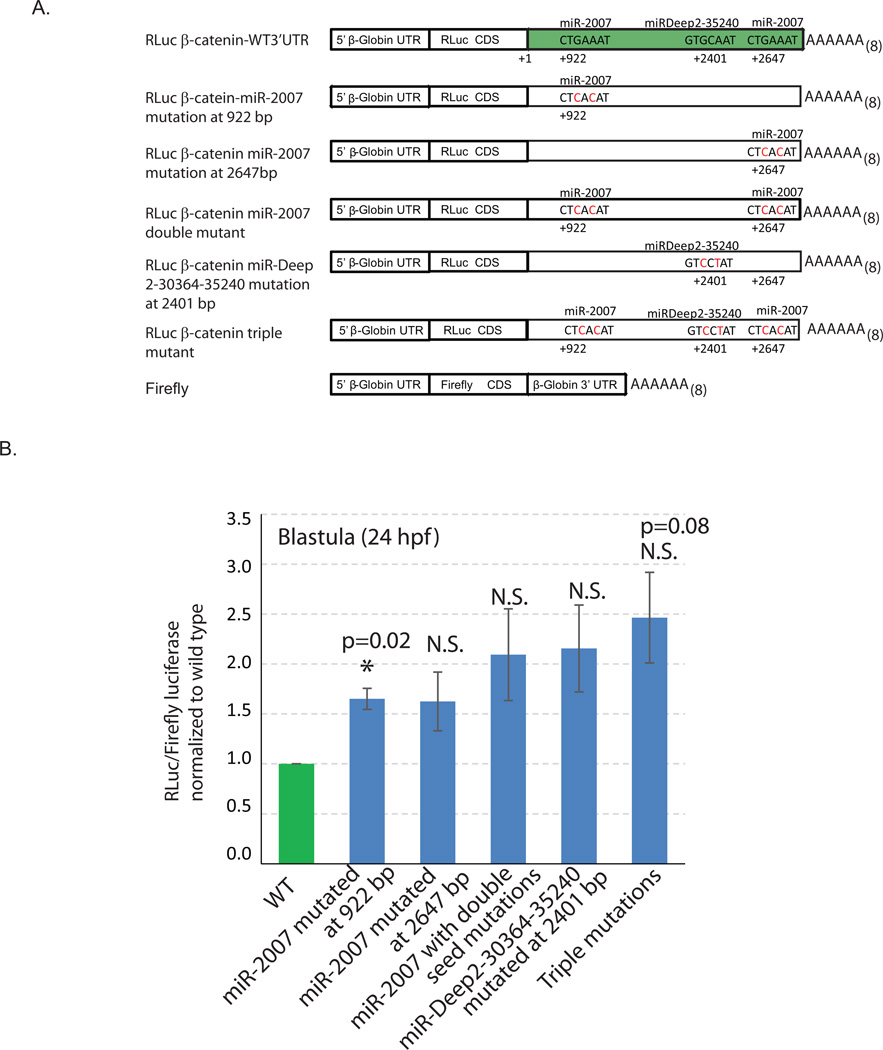
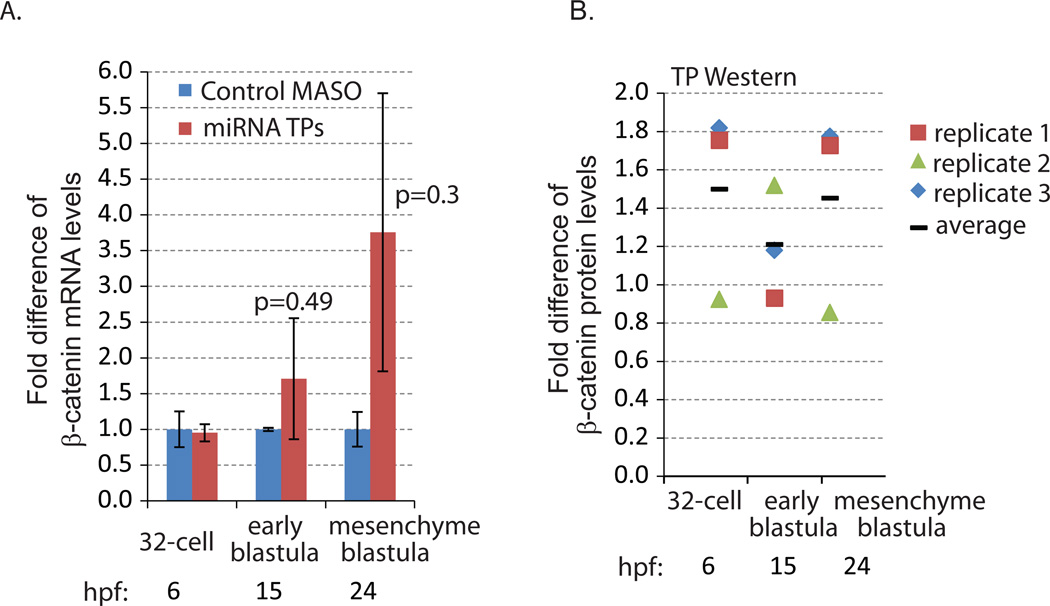
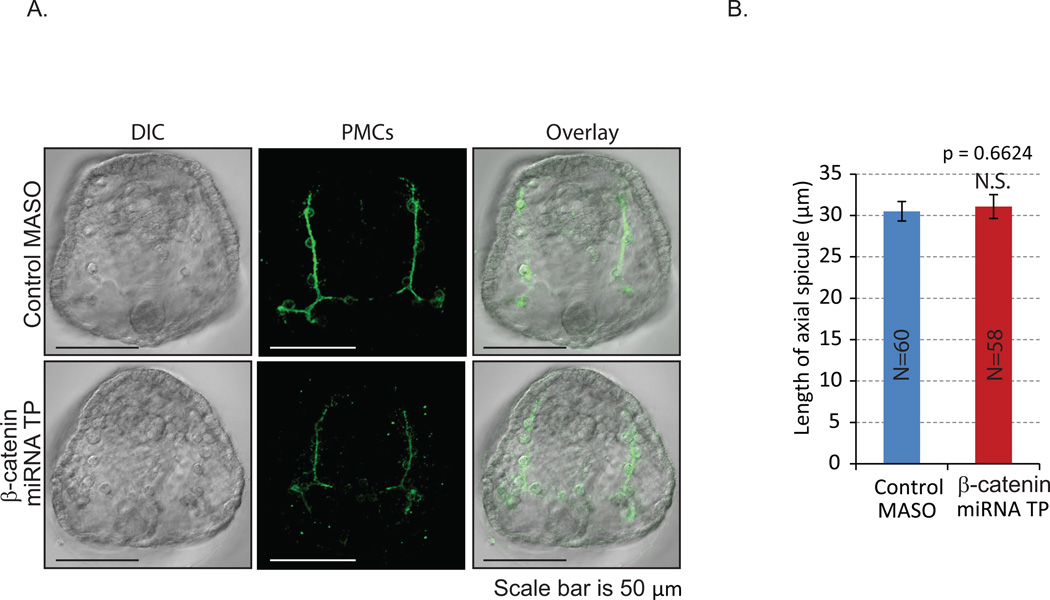
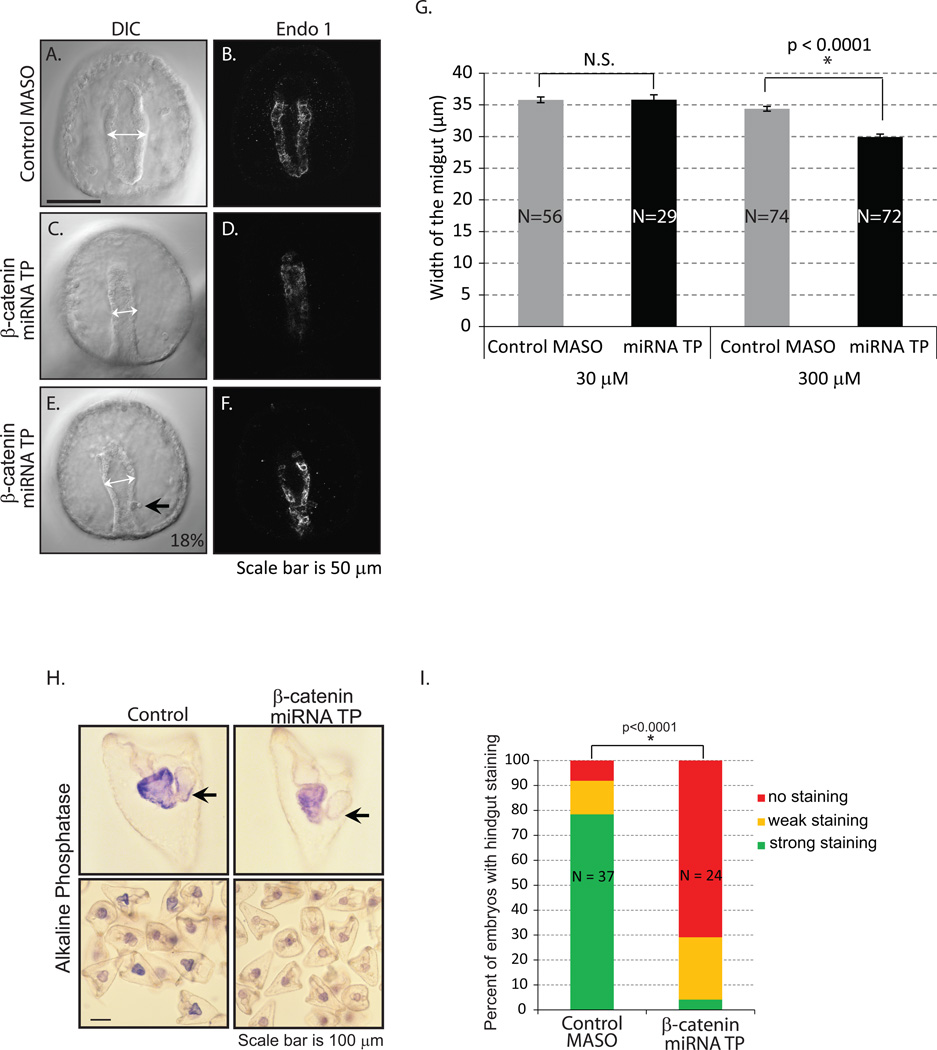
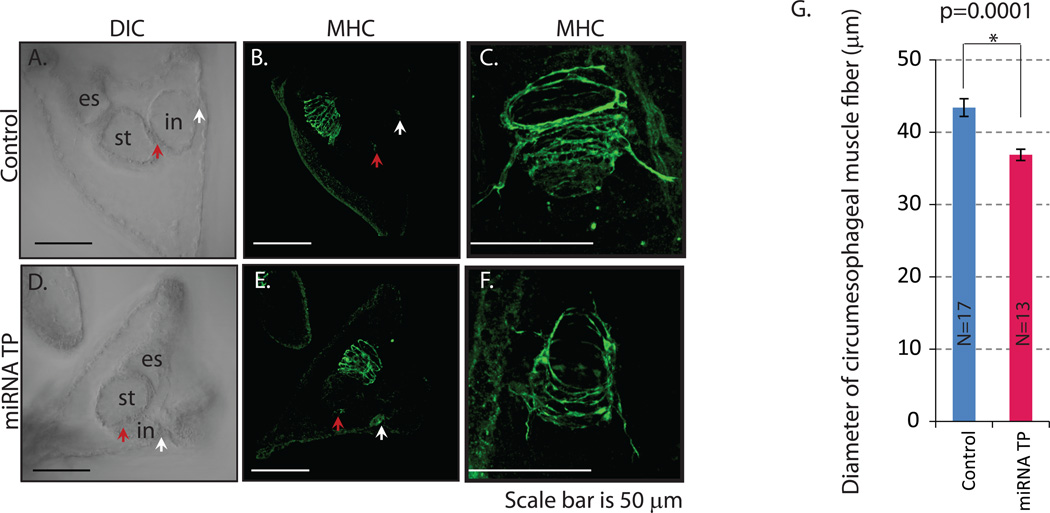
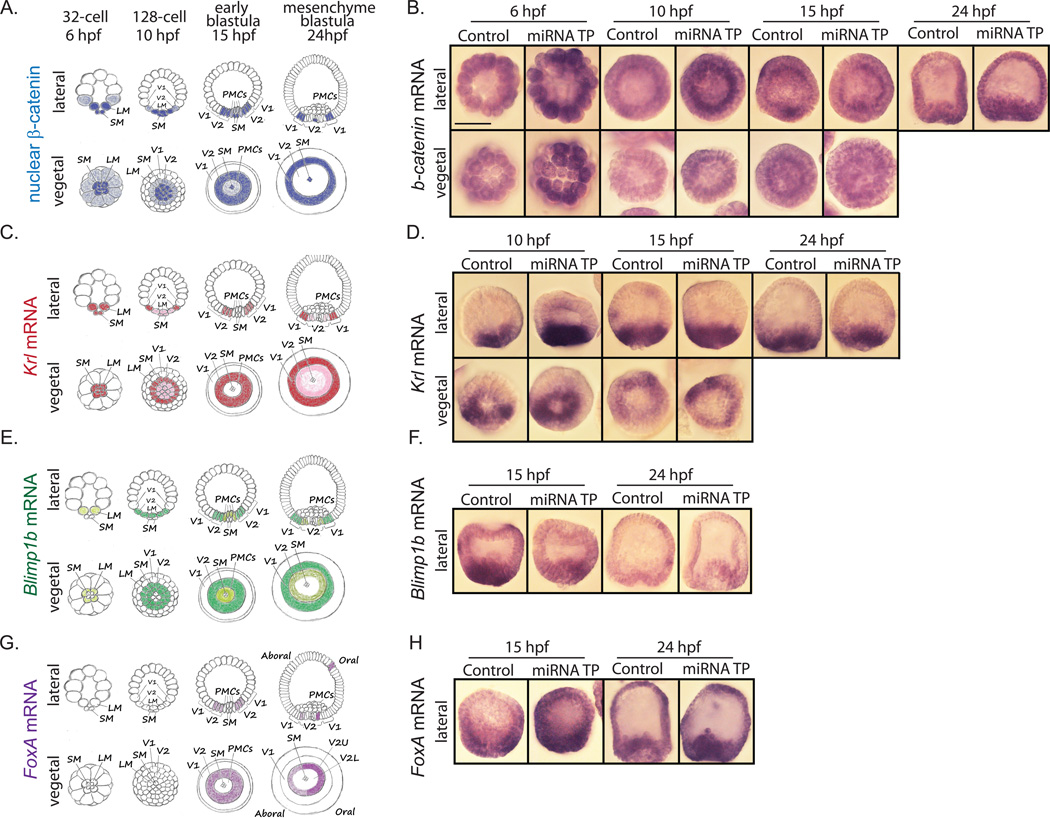
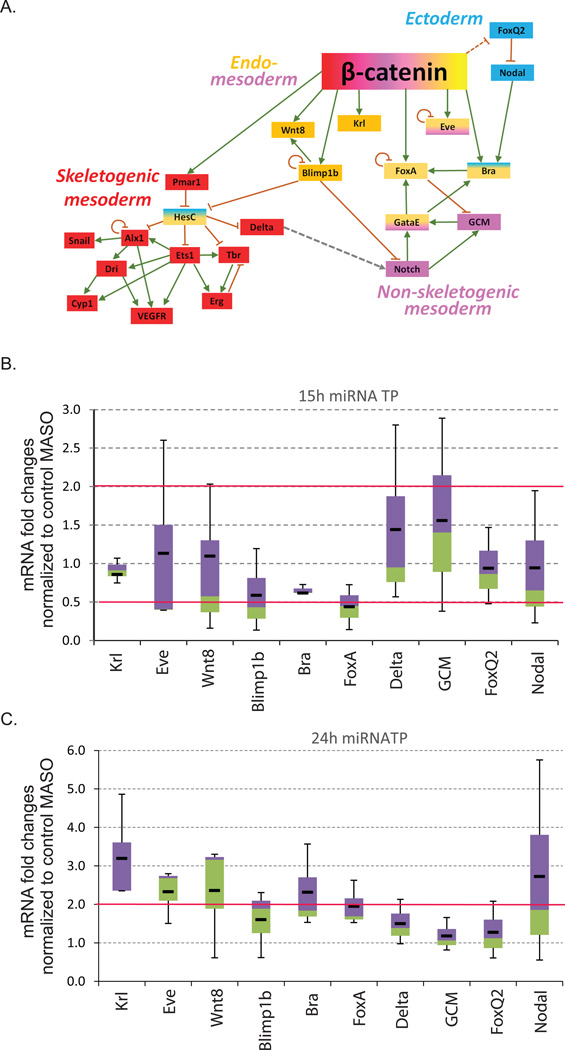
Development of complex multicellular organisms requires careful regulation at both transcriptional and post-transcriptional levels. Post-transcriptional gene regulation is in part mediated by a class of non-coding RNAs of 21-25 nucleotides in length known as microRNAs (miRNAs). β-catenin, regulated by the canonical Wnt signaling pathway, has a highly evolutionarily conserved function in patterning early metazoan embryos, in forming the Anterior-Posterior axis, and in establishing the endomesoderm. Using reporter constructs and site-directed mutagenesis, we identified at least three miRNA binding sites within the 3' untranslated region (3'UTR) of the sea urchin β-catenin. Further, blocking these three miRNA binding sites within the β-catenin 3'UTR to prevent regulation of endogenous β-catenin by miRNAs resulted in a minor increase in β-catenin protein accumulation that is sufficient to induce aberrant gut morphology and circumesophageal musculature. These phenotypes are likely the result of increased transcript levels of Wnt responsive endomesodermal regulatory genes. This study demonstrates the importance of miRNA regulation of β-catenin in early development.
Keywords: Circumesophageal muscles; Endoderm; Larval gut; Mesoderm; PMCs; Post-transcriptional regulation; Sea urchin; Target protectors.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
References
-
- Aberle H, Schwartz H, Kemler R. Cadherin-catenin complex: protein interactions and their implications for cadherin function. J Cell Biochem. 1996;61:514–523. - PubMed
-
- Anastas JN, Moon RT. WNT signalling pathways as therapeutic targets in cancer. Nat Rev Cancer. 2013;13:11–26. - PubMed
-
- Annunziata R, Perillo M, Andrikou C, Cole AG, Martinez P, Arnone MI. Pattern and process during sea urchin gut morphogenesis: The regulatory landscape. Genesis. 2013 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
