

Review
doi: 10.1038/nri3800.
Role of innate and adaptive immune mechanisms in cardiac injury and repair
Affiliations
- PMID: 25614321
- PMCID: PMC4669103
- DOI: 10.1038/nri3800
Item in Clipboard
Review
Role of innate and adaptive immune mechanisms in cardiac injury and repair
Nat Rev Immunol.
2015 Feb.
Abstract
Despite the advances that have been made in developing new therapeutics, cardiovascular disease remains the leading cause of worldwide mortality. Therefore, understanding the mechanisms underlying cardiovascular tissue injury and repair is of prime importance. Following cardiac tissue injury, the immune system has an important and complex role in driving both the acute inflammatory response and the regenerative response. This Review summarizes the role of the immune system in cardiovascular disease - focusing on the idea that the immune system evolved to promote tissue homeostasis following injury and/or infection, and that the inherent cost of this evolutionary development is unwanted inflammatory damage.
Figures
Schematic demonstrating coronary artery occlusion (black) that leads to ischemic tissue injury (grey zone). From within the ischemic zone, cell necrosis, extracellular matrix (ECM) degradation and recruitment of immune cells all leads to production specific damage associated molecular patterns (DAMP)s, which are recognized by pattern recognition receptors (PRR)s, leading to the generation of inflammatory responses to internal injury signals.
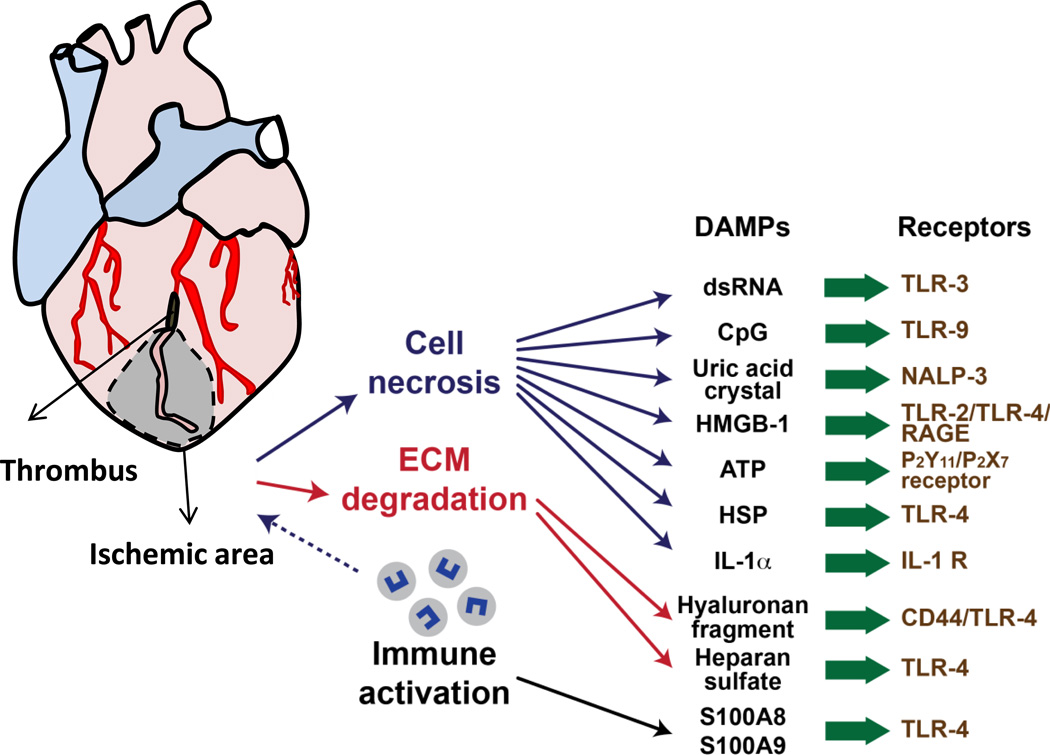
Schematic demonstrating coronary artery occlusion (black) that leads to ischemic tissue injury (grey zone). From within the ischemic zone, cell necrosis, extracellular matrix (ECM) degradation and recruitment of immune cells all leads to production specific damage associated molecular patterns (DAMP)s, which are recognized by pattern recognition receptors (PRR)s, leading to the generation of inflammatory responses to internal injury signals.
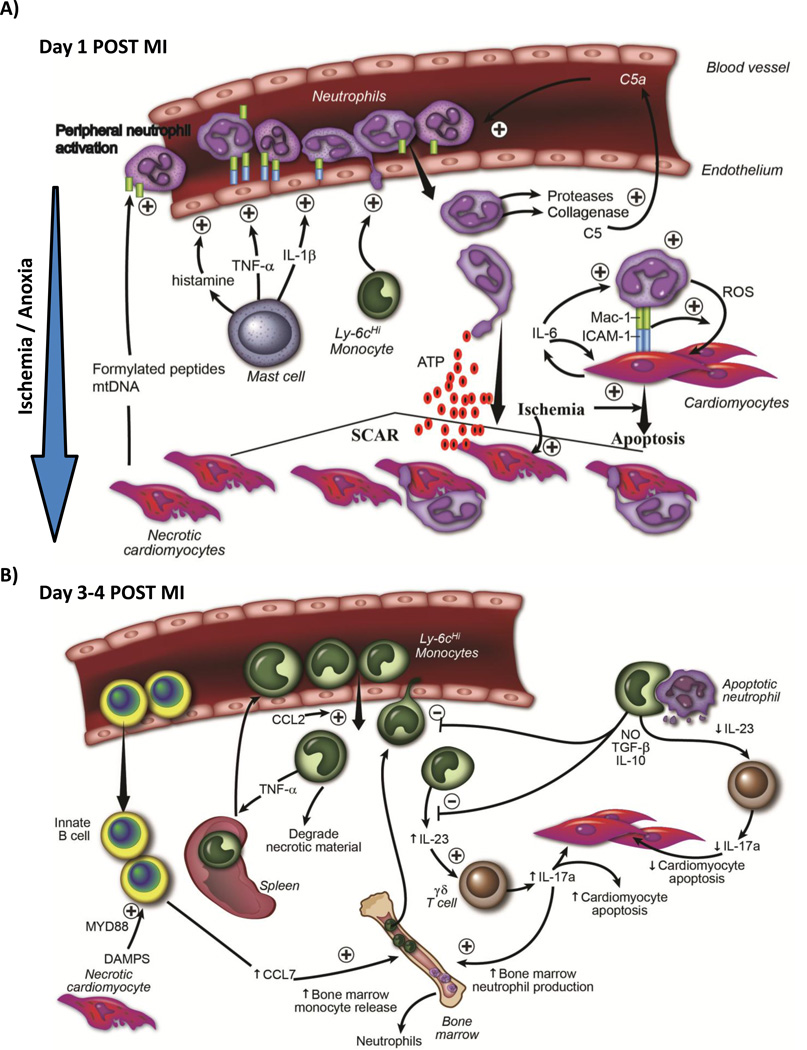
A) Temporal schematic demonstrating that early after ischemic injury (first 24 hrs), internal DAMP signals released from necrotic cardiomyocytes activate resident mast cells, causing degranulation and release of preformed pro-inflammatory cytokines and vasogenic compounds such as histamine, which activate endothelial cells. Necrotic cardiomyocytes also release mitochondrial DAMPS (formylated peptides and mtDNA) into circulation, which causes systemic neutrophil activation. Activated neutrophils adhere to activated endothelium, transmigrate into tissue following a chemokine gradient. Neutrophils secrete proteases that digest tissue (and also activate chemokines such as C5a), which further potentiates leukocyte recruitment. Early recruitment of monocytes aids neutrophil recruitment. Neutrophils are directed to ischemic areas by following DAMP gradient (such as ATP). Neutrophils then both phagocytose dying cells, but can also induce apoptosis in live cardiomyocytes themselves through a MAC-1 : ICAM-1 interaction and release of reactive oxygen species. B) Schematic demonstrating that later after ischemic injury (24-96 hrs), there is recruitment of Ly6cHi monocytes from the blood into ischemic cardiac tissue. Some of the monocytes originated from the spleen. IgM/IgD+ innate B cells are also recruited into the myocardium, and through a CCL7-dependent fashion, promote further monocyte recruitment . Innate B cell activation is Myd88 dependent (suggesting TLR / DAMP involvement). Recruited monocytes secrete pro-inflammatory cytokines and chemokines, and drive inflammatory processes. A proportion of recruited monocytes ingested apoptotic material including neutrophils, which serves to increase secretion of anti-inflammatory cytokines such as TGF-B and IL-10, and thereby decrease leukocyte recruitment. Monocytes produce IL-23, which drives innate γδ T cells to produce IL-17a. IL-17a has two roles; it drives neutrophil production in the bone marrow and causes cardiomyocytes death. As inflammatory responses diminish, less IL-23 is produced.
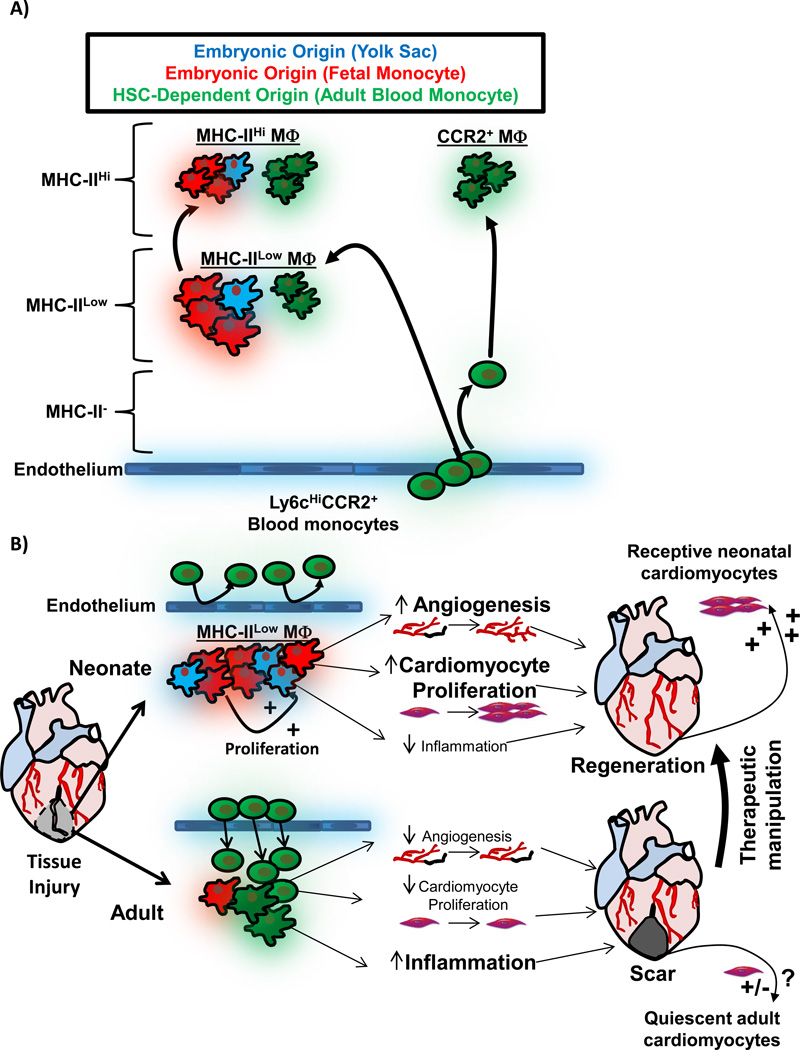
A) Schematic depicting cardiac MΦ origins. During embryonic development, initially extra-embryonic yolk-sac derived MΦs seed the developing heart (blue), and expressed low levels of MHC-II. Later during development, fetal-monocyte-derived MΦs also infiltrate the heart (red), and both embryonic MΦ populations expand by proliferation during development and after birth. MHC-IIHi MΦs develop from MHC-IILow MΦs after weaning , . During this time, HSC-derived monocytes infiltrate the heart and also differentiate into MHC-IIHi and MHC-IILow MΦs (green). MHC-IIHi and MHC-IILow MΦ live as ontologically mixed groups, primarily made up of embryonic-derived MΦs . Monocytes infiltrate the heart and become shorter lived CCR2+ MΦs, which are entirely derived from blood monocytes . B) Comparison of the MΦ response to tissue injury in neonatal animals (which regenerate cardiac tissue) and adult animals (minimal regeneration). In the neonate, resident embryonic-derived populations expand without significant monocyte input. Embryonic-derived MΦs promote angiogenesis, cardiomyocyte proliferation and produce minimal inflammation when stimulated by DAMPs, which together facilitate cardiac regeneration . In injury in the adult, large numbers expansion of CCR2+ monocytes and MΦs, which possess a limited ability to promote angiogenesis and cardiomyocyte expansion, but have significant capacity to drive inflammatory responses, which impedes regeneration , . Importantly, neonatal cardiomyocyte are primed to divide and therefore "receptive" to regenerative signals , , . Adult cardiomyocytes are much less receptive to regenerative signals, and growth signals from embryonic-derived cardiac MΦs are but one important component to enhancing cardiac regeneration in adult tissues. To exploit this understanding therapeutically, the goal would be model the neonatal response to injury (embryonic-derived MΦs and receptive cardiomyocytes) in the adult.
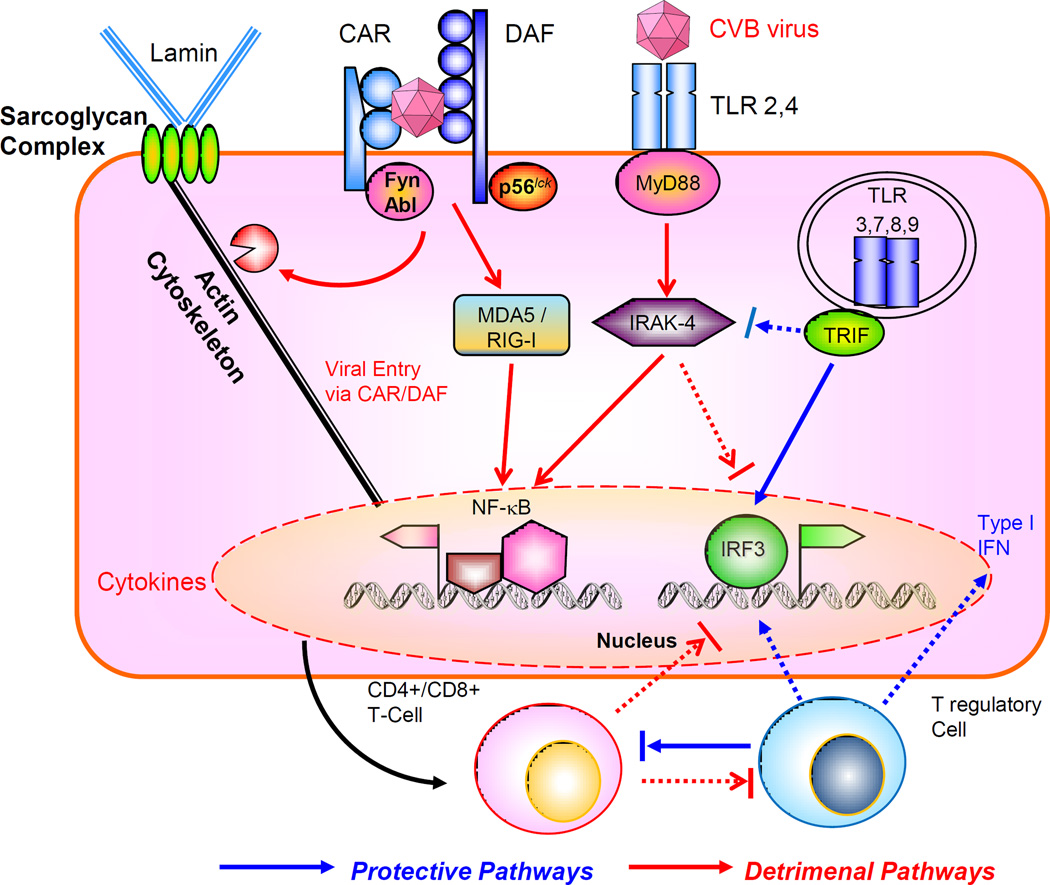
The virus engages the internalizing receptor Coxsackie-Adeno Receptor (CAR) whose tyrosine kinases Fyn and Abl can facilitate viral remodeling of cytoskeleton to gain entry, further aided by co-receptor decay accelerating factor (DAF), associated with tyrosine kinase p56lck. The viral components can interact with MDA5/RIG-I to activate NF-kB, or engage the cell surface toll-like receptors (TLR) through adaptor MyD88 and downstream signal intermediates such as IRAK-4 and TRAF6. These pathways facilitate viral proliferation and host immune tissue damage. On the other hand, activation of TRIF-IRF3/IRF7 pathway leading to type I interferon (IFN) production is protective. There is mutual counter-regulation of the MyD88 –IRAK4 and TRIF-IRF3 pathways. The subsequent maturation of CD4+ / CD8+ T cell subsets is also detrimental for the host, in contrast to the T regulatory cells which are host-protective. Cross talk between acquired and innate immune signaling pathways modulate the host response to viral infection.
References
-
- Bier E, Bodmer R. Drosophila, an emerging model for cardiac disease. Gene. 2004;342:1–11. - PubMed
-
- Poss KD, Wilson LG, Keating MT. Heart regeneration in zebrafish. Science. 2002;298:2188–2190. - PubMed
-
- Laube F, Heister M, Scholz C, Borchardt T, Braun T. Re-programming of newt cardiomyocytes is induced by tissue regeneration. J. Cell Sci. 2006;119:4719–4729. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
