

Soybean kinome: functional classification and gene expression patterns
- PMID: 25614662
- PMCID: PMC4378628
- DOI: 10.1093/jxb/eru537
Soybean kinome: functional classification and gene expression patterns
Abstract
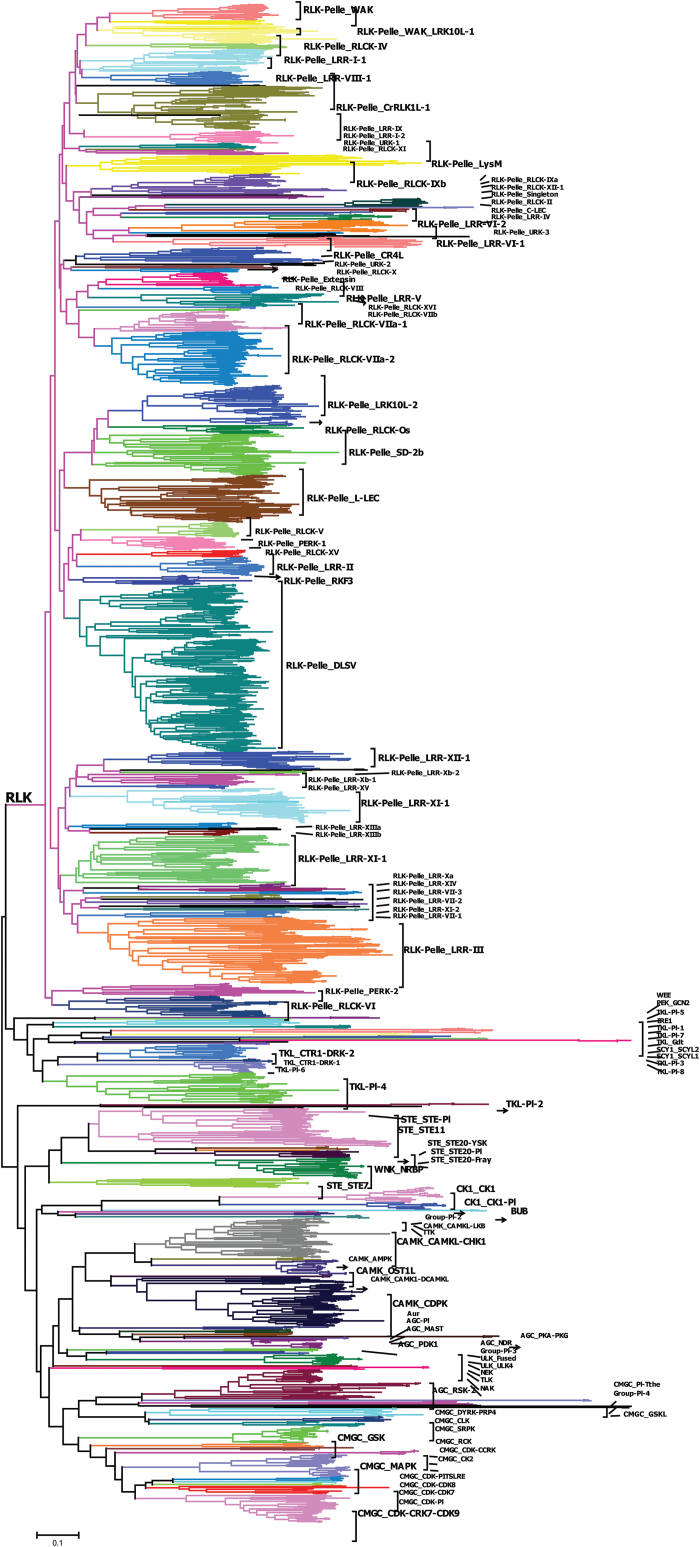
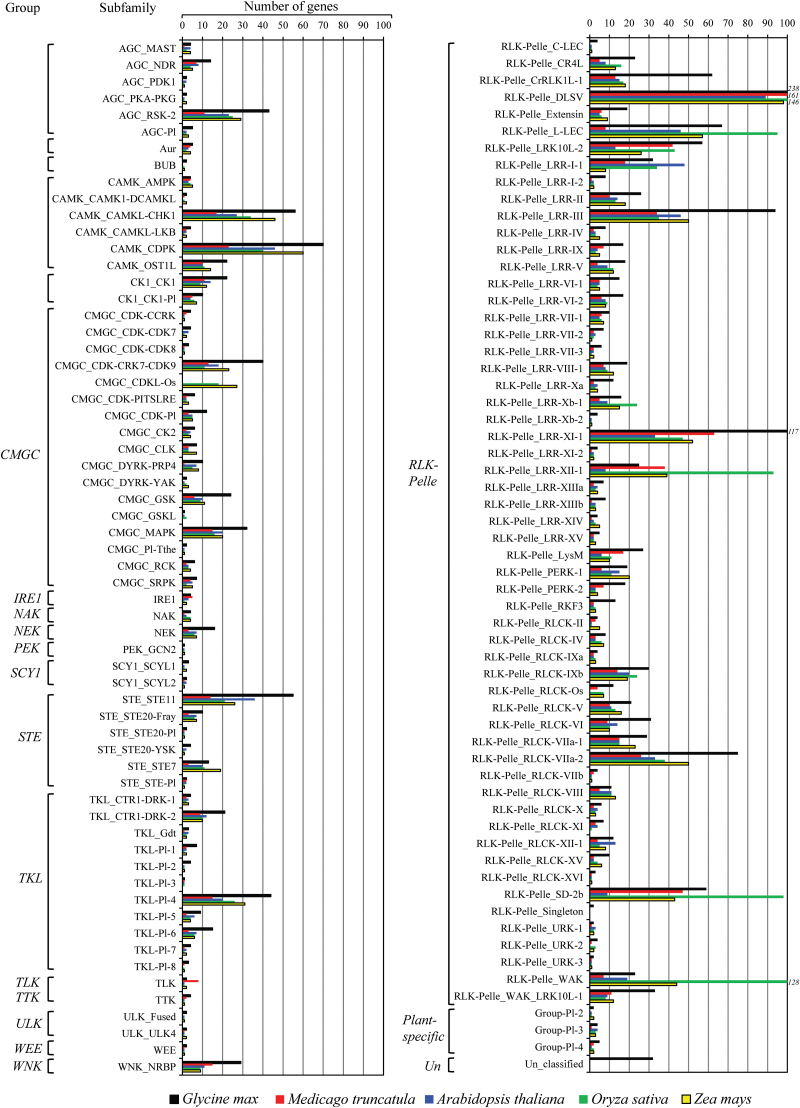
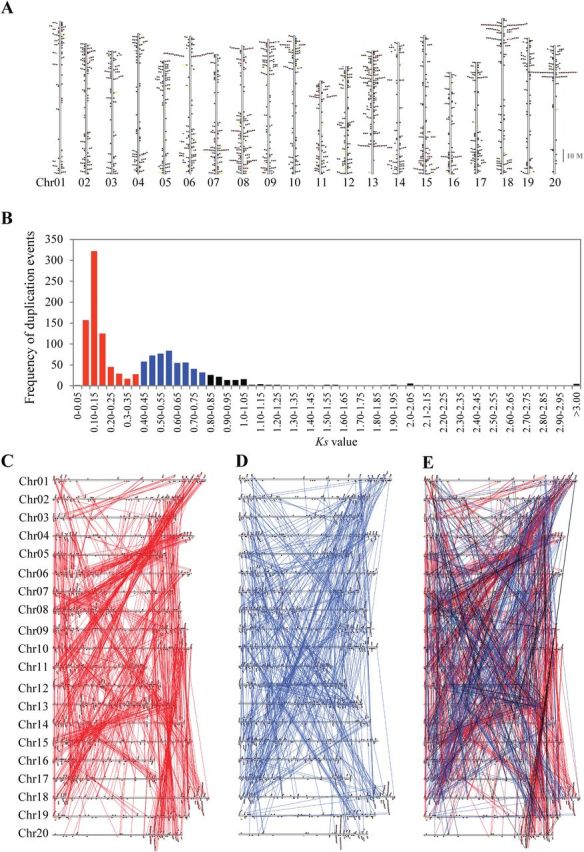
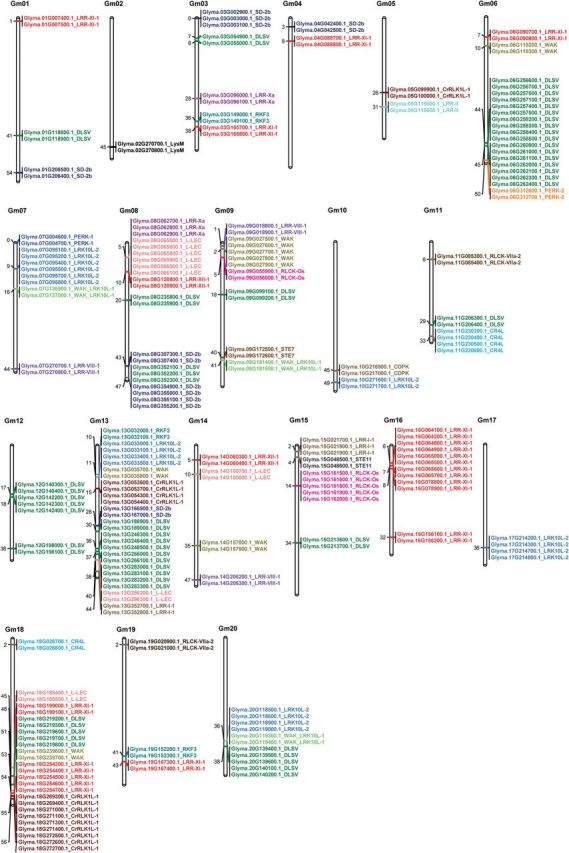
The protein kinase (PK) gene family is one of the largest and most highly conserved gene families in plants and plays a role in nearly all biological functions. While a large number of genes have been predicted to encode PKs in soybean, a comprehensive functional classification and global analysis of expression patterns of this large gene family is lacking. In this study, we identified the entire soybean PK repertoire or kinome, which comprised 2166 putative PK genes, representing 4.67% of all soybean protein-coding genes. The soybean kinome was classified into 19 groups, 81 families, and 122 subfamilies. The receptor-like kinase (RLK) group was remarkably large, containing 1418 genes. Collinearity analysis indicated that whole-genome segmental duplication events may have played a key role in the expansion of the soybean kinome, whereas tandem duplications might have contributed to the expansion of specific subfamilies. Gene structure, subcellular localization prediction, and gene expression patterns indicated extensive functional divergence of PK subfamilies. Global gene expression analysis of soybean PK subfamilies revealed tissue- and stress-specific expression patterns, implying regulatory functions over a wide range of developmental and physiological processes. In addition, tissue and stress co-expression network analysis uncovered specific subfamilies with narrow or wide interconnected relationships, indicative of their association with particular or broad signalling pathways, respectively. Taken together, our analyses provide a foundation for further functional studies to reveal the biological and molecular functions of PKs in soybean.
Keywords: abiotic stress; biotic stress; co-expression network; collinearity analysis; duplication events; gene expression; soybean kinases..
© The Author 2015. Published by Oxford University Press on behalf of the Society for Experimental Biology.
Figures
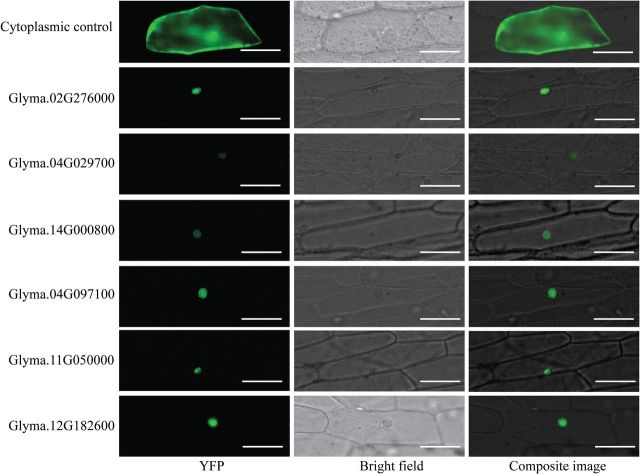
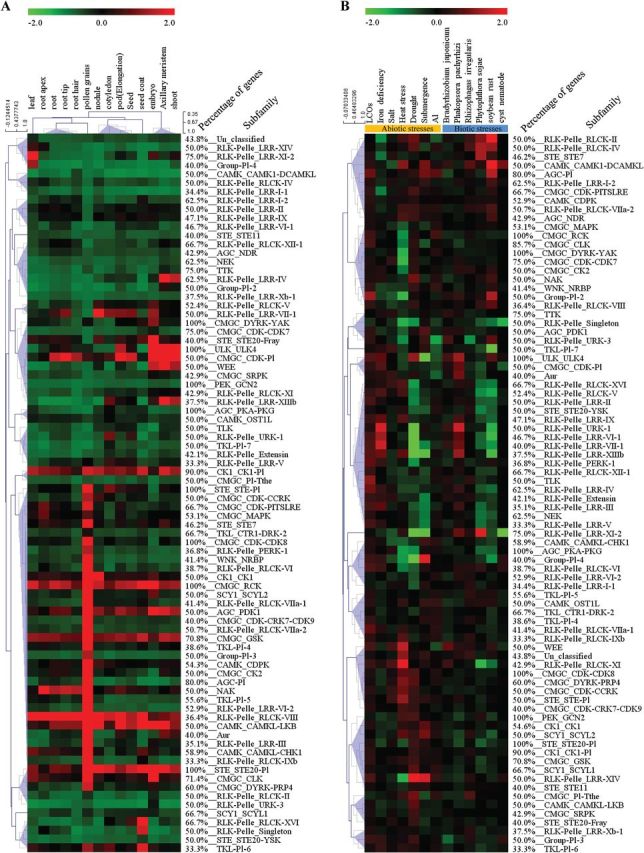
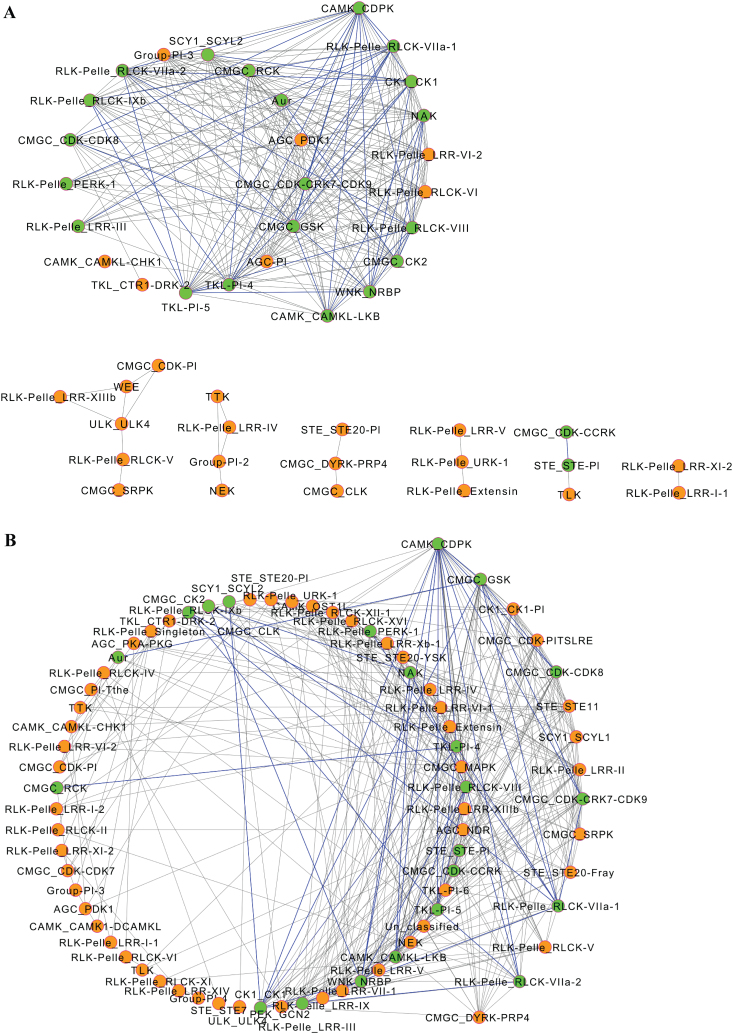
References
-
- Antolín-Llovera M, Ried MK, Binder A, Parniske M. 2012. Receptor kinase signaling pathways in plant–microbe interactions. Annual Review of Phytopathology 50, 451–473. - PubMed
-
- Brameier M, Krings A, MacCallum RM. 2007. NucPred—predicting nuclear localization of proteins. Bioinformatics 23, 1159–1160. - PubMed
-
- Champion A, Kreis M, Mockaitis K, Picaud A, Henry Y. 2004. Arabidopsis kinome: after the casting. Functional and Integrative Genomics 4, 163–187. - PubMed
-
- Chern M, Fitzgerald HA, Canlas PE, Navarre DA, Ronald PC. 2005. Overexpression of a rice NPR1 homolog leads to constitutive activation of defense response and hypersensitivity to light. Molecular Plant–Microbe Interactions 18, 511–520. - PubMed
-
- Colcombet J, Hirt H. 2008. Arabidopsis MAPKs: a complex signalling network involved in multiple biological processes. Biochemical Journal 413, 217–226. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
