

HLA-A02:01-restricted epitopes identified from the herpes simplex virus tegument protein VP11/12 preferentially recall polyfunctional effector memory CD8+ T cells from seropositive asymptomatic individuals and protect humanized HLA-A*02:01 transgenic mice against ocular herpes
- PMID: 25617474
- PMCID: PMC4340763
- DOI: 10.4049/jimmunol.1402606
HLA-A02:01-restricted epitopes identified from the herpes simplex virus tegument protein VP11/12 preferentially recall polyfunctional effector memory CD8+ T cells from seropositive asymptomatic individuals and protect humanized HLA-A*02:01 transgenic mice against ocular herpes
Abstract
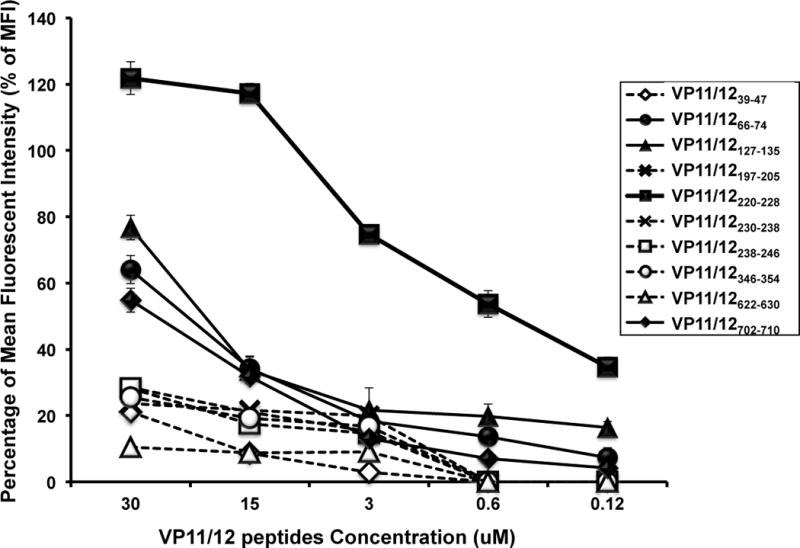
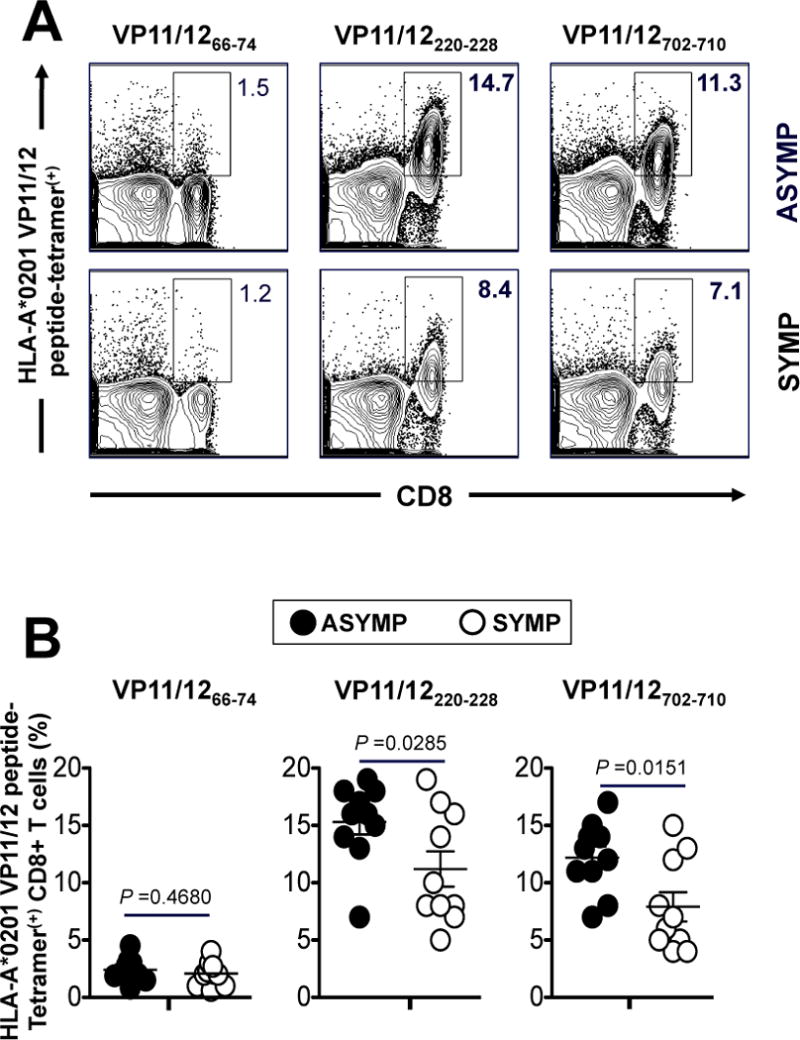
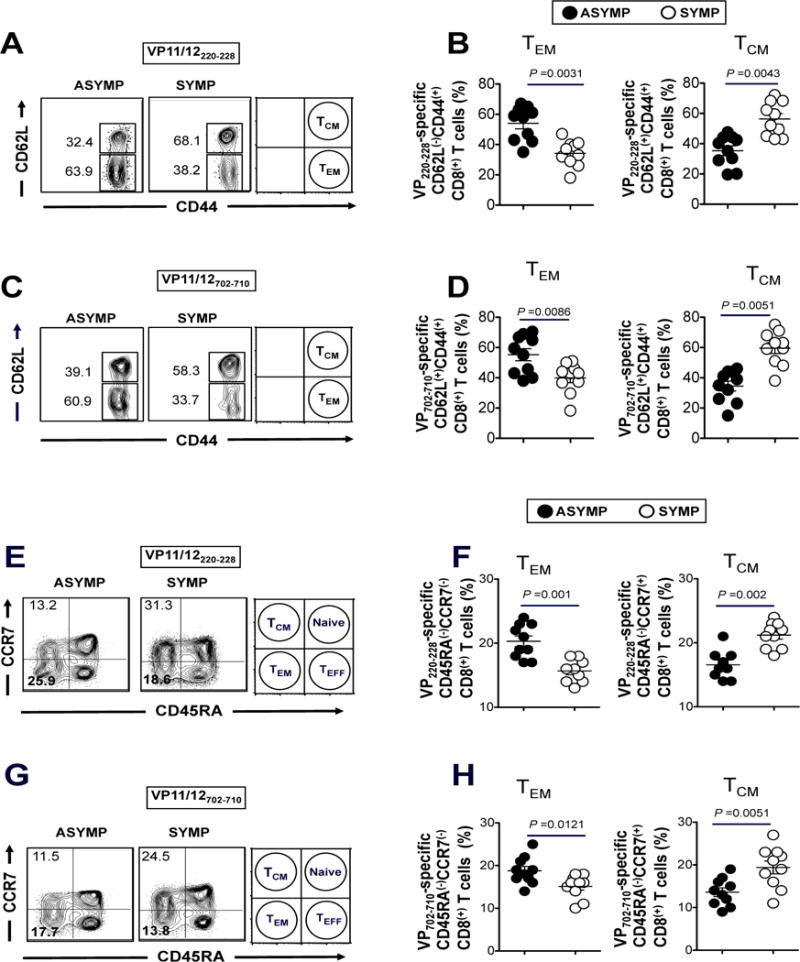
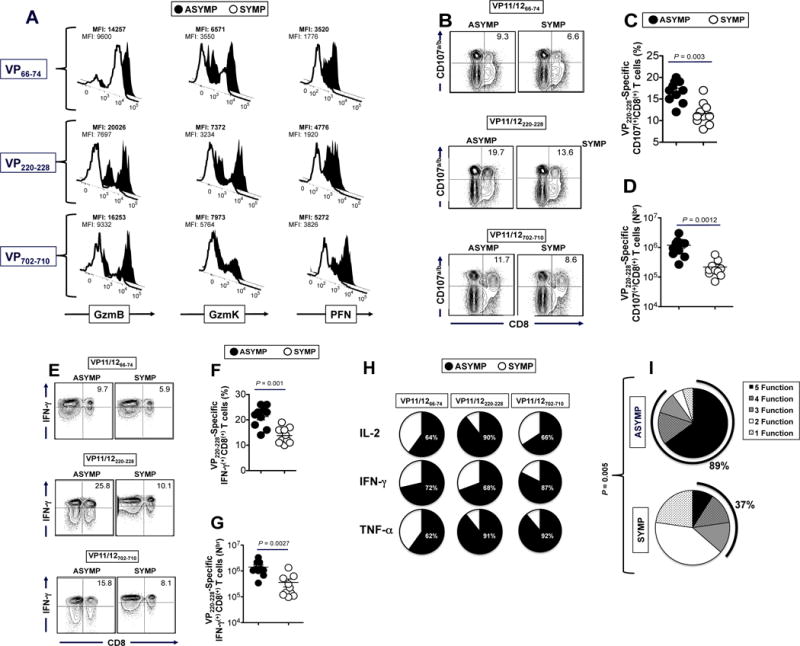
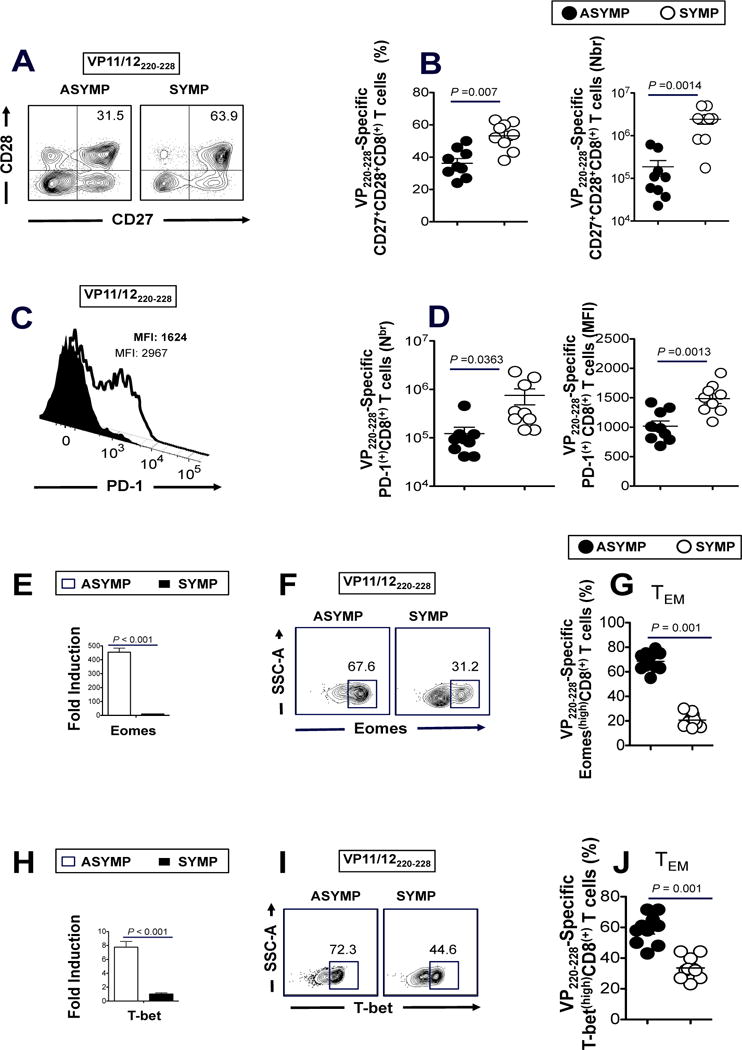
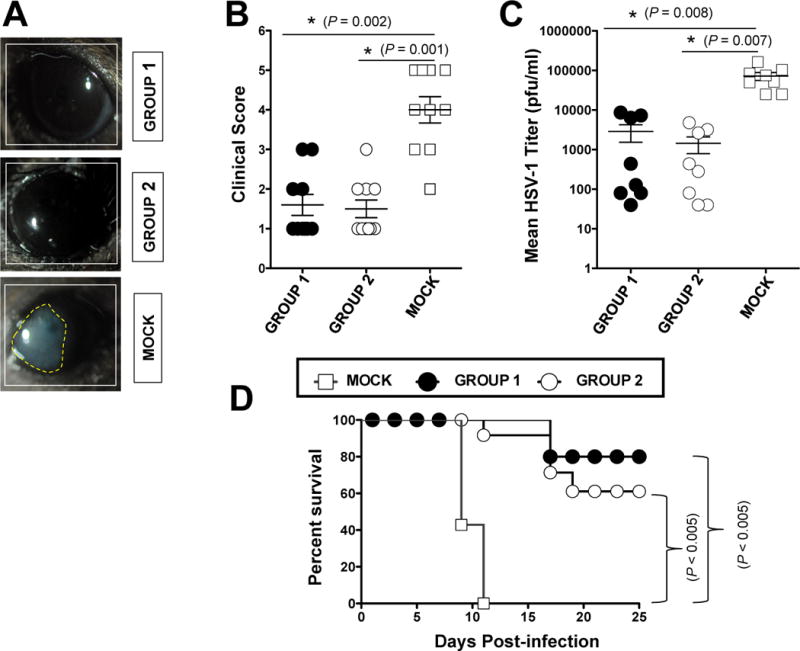
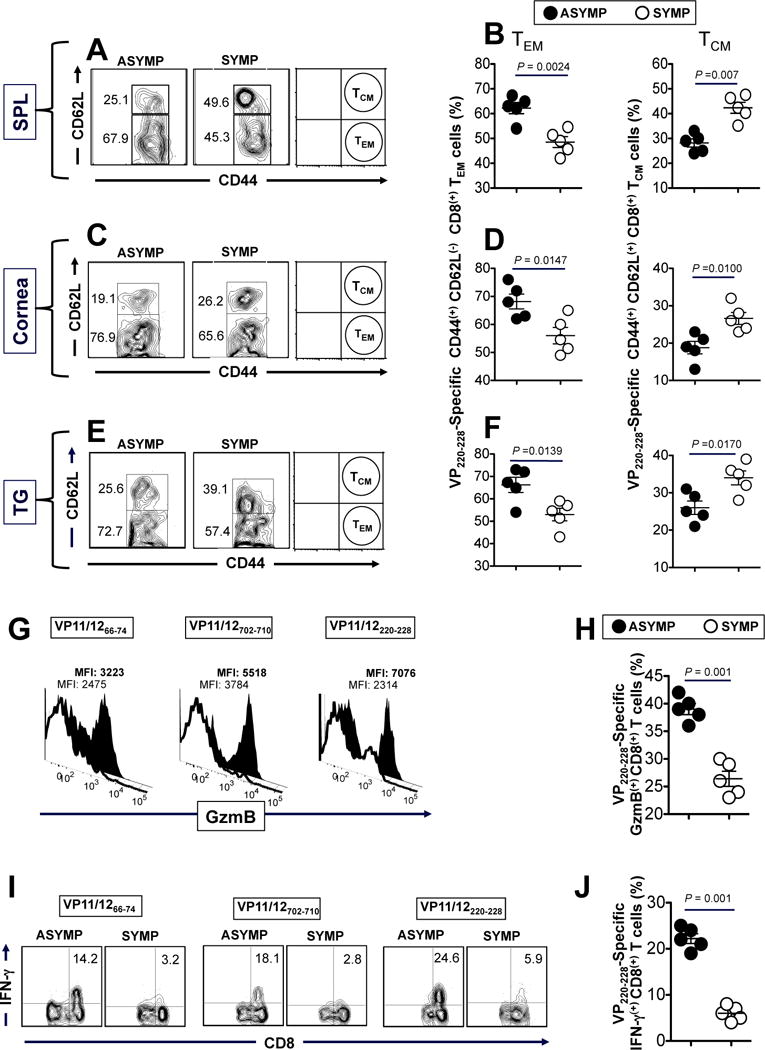
The HSV type 1 tegument virion phosphoprotein (VP) 11/12 (VP11/12) is a major Ag targeted by CD8(+) T cells from HSV-seropositive individuals. However, whether and which VP11/12 epitope-specific CD8(+) T cells play a role in the "natural" protection seen in seropositive healthy asymptomatic (ASYMP) individuals (who have never had clinical herpes disease) remain to be determined. In this study, we used multiple prediction computer-assisted algorithms to identify 10 potential HLA-A*02:01-restricted CD8(+) T cell epitopes from the 718-aa sequence of VP11/12. Three of 10 epitopes exhibited high-to-moderate binding affinity to HLA-A*02:01 molecules. In 10 sequentially studied HLA-A*02:01-positive and HSV-1-seropositive ASYMP individuals, the most frequent, robust, and polyfunctional effector CD8(+) T cell responses, as assessed by a combination of tetramer frequency, granzyme B, granzyme K, perforin, CD107(a/b) cytotoxic degranulation, IFN-γ, and multiplex cytokines assays, were predominantly directed against three epitopes: VP11/1266-74, VP11/12220-228, and VP11/12702-710. Interestingly, ASYMP individuals had a significantly higher proportion of CD45RA(low)CCR7(low)CD44(high)CD62L(low)CD27(low)CD28(low)CD8(+) effector memory CD8(+) T cells (TEMs) specific to the three epitopes, compared with symptomatic individuals (with a history of numerous episodes of recurrent ocular herpetic disease). Moreover, immunization of HLA-A*02:01 transgenic mice with the three ASYMP CD8(+) TEM cell epitopes induced robust and polyfunctional epitope-specific CD8(+) TEM cells that were associated with a strong protective immunity against ocular herpes infection and disease. Our findings outline phenotypic and functional features of protective HSV-specific CD8(+) T cells that should guide the development of an effective T cell-based herpes vaccine.
Copyright © 2015 by The American Association of Immunologists, Inc.
Conflict of interest statement
Conflict of interest: The authors have declared that no conflict of interest exists
Figures
References
-
- Dervillez X, Qureshi H, Chentoufi AA, Khan AA, Kritzer E, Yu DC, Diaz OR, Gottimukkala C, Kalantari M, Villacres MC, Scarfone VM, McKinney DM, Sidney J, Sette A, Nesburn AB, Wechsler SL, BenMohamed L. Asymptomatic HLA-A*02:01-restricted epitopes from herpes simplex virus glycoprotein B preferentially recall polyfunctional CD8+ T cells from seropositive asymptomatic individuals and protect HLA transgenic mice against ocular herpes. J Immunol. 2013;191:5124–5138. - PMC - PubMed
-
- Chentoufi AA, Zhang X, Lamberth K, Dasgupta G, Bettahi I, Nguyen A, Wu M, Zhu X, Mohebbi A, Buus S, Wechsler SL, Nesburn AB, BenMohamed L. HLA-A*0201-restricted CD8+ cytotoxic T lymphocyte epitopes identified from herpes simplex virus glycoprotein D. J Immunol. 2008;180:426–437. - PubMed
-
- Zhang X, Dervillez X, Chentoufi AA, Badakhshan T, Bettahi I, Benmohamed L. Targeting the genital tract mucosa with a lipopeptide/recombinant adenovirus prime/boost vaccine induces potent and long-lasting CD8+ T cell immunity against herpes: importance of MyD88. J Immunol. 2012;189:4496–4509. - PMC - PubMed
-
- Samandary S, Kridane-Miledi H, Sandoval JS, Choudhury Z, Langa-Vives F, Spencer D, Chentoufi AA, Lemonnier FA, BenMohamed L. Associations of HLA-A, HLA-B and HLA-C alleles frequency with prevalence of herpes simplex virus infections and diseases across global populations: implication for the development of an universal CD8+ T-cell epitope-based vaccine. Hum Immunol. 2014;75:715–729. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
