

The molecular basis for histone H4- and H2A-specific amino-terminal acetylation by NatD
- PMID: 25619998
- PMCID: PMC4318724
- DOI: 10.1016/j.str.2014.10.025
The molecular basis for histone H4- and H2A-specific amino-terminal acetylation by NatD
Abstract
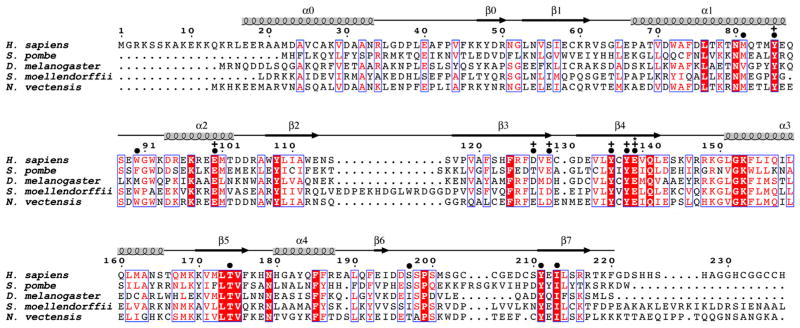
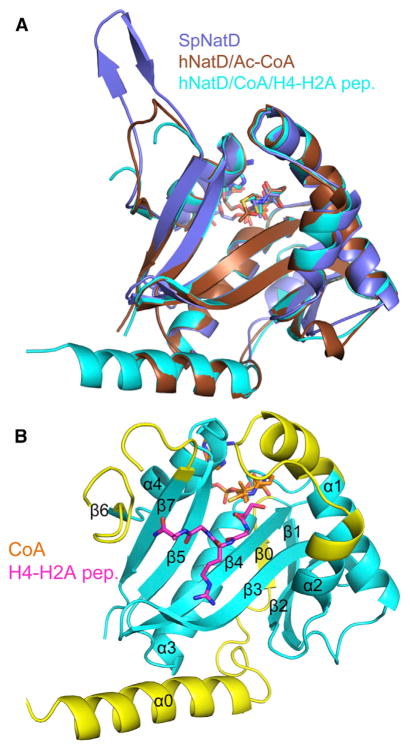
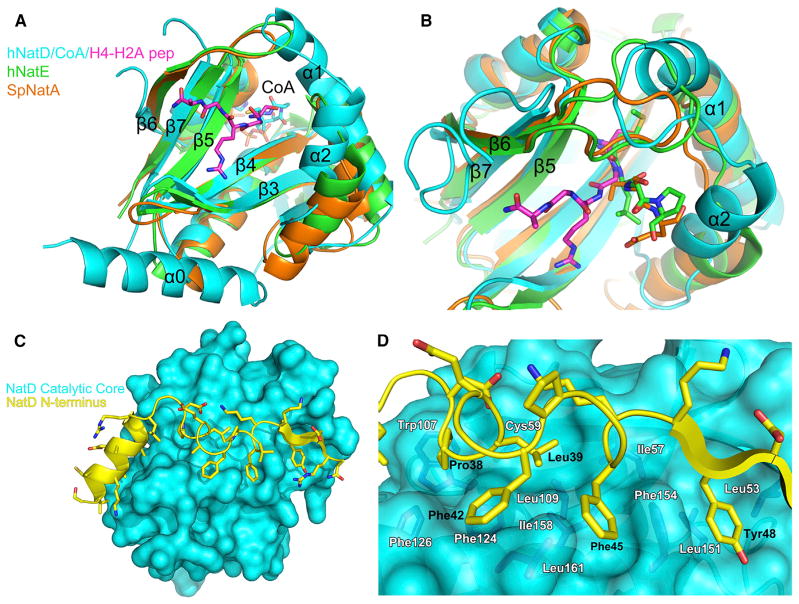
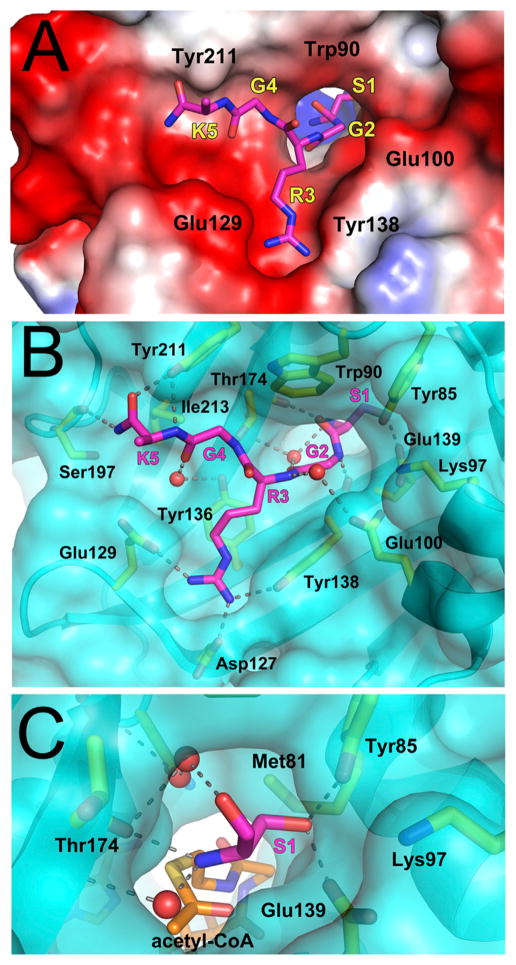
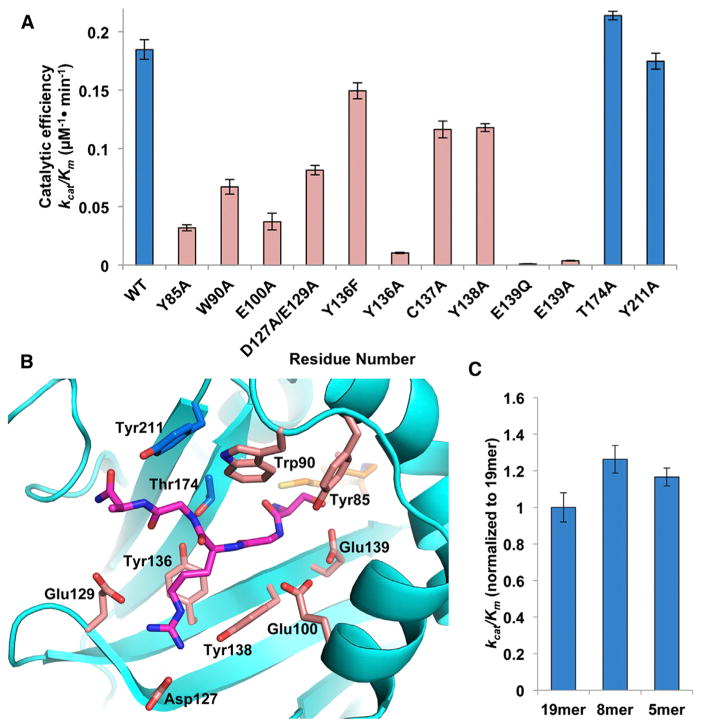
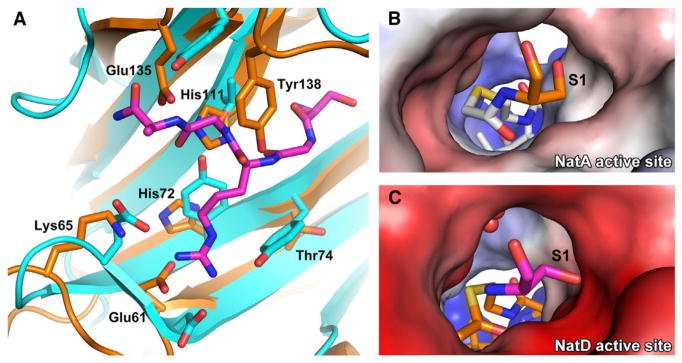
N-terminal acetylation is among the most common protein modifications in eukaryotes and is mediated by evolutionarily conserved N-terminal acetyltransferases (NATs). NatD is among the most selective NATs; its only known substrates are histones H4 and H2A, containing the N-terminal sequence SGRGK in humans. Here we characterize the molecular basis for substrate-specific acetylation by NatD by reporting its crystal structure bound to cognate substrates and performing related biochemical studies. A novel N-terminal segment wraps around the catalytic core domain to make stabilizing interactions, and the α1-α2 and β6-β7 loops adopt novel conformations to properly orient the histone N termini in the binding site. Ser1 and Arg3 of the histone make extensive contacts to highly conserved NatD residues in the substrate binding pocket, and flanking glycine residues also appear to contribute to substrate-specific binding by NatD, together defining a Ser-Gly-Arg-Gly recognition sequence. These studies have implications for understanding substrate-specific acetylation by NAT enzymes.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
