

Paths to a malaria vaccine illuminated by parasite genomics
- PMID: 25620796
- PMCID: PMC4359294
- DOI: 10.1016/j.tig.2014.12.005
Paths to a malaria vaccine illuminated by parasite genomics
Abstract
More human death and disease is caused by malaria parasites than by all other eukaryotic pathogens combined. As early as the sequencing of the first human genome, malaria parasite genomics was prioritized to fuel the discovery of vaccine candidate antigens. This stimulated increased research on malaria, generating new understanding of the cellular and molecular mechanisms of infection and immunity. This review of recent developments illustrates how new approaches in parasite genomics, and increasingly large amounts of data from population studies, are helping to identify antigens that are promising lead targets. Although these results have been encouraging, effective discovery and characterization need to be coupled with more innovation and funding to translate findings into newly designed vaccine products for clinical trials.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures
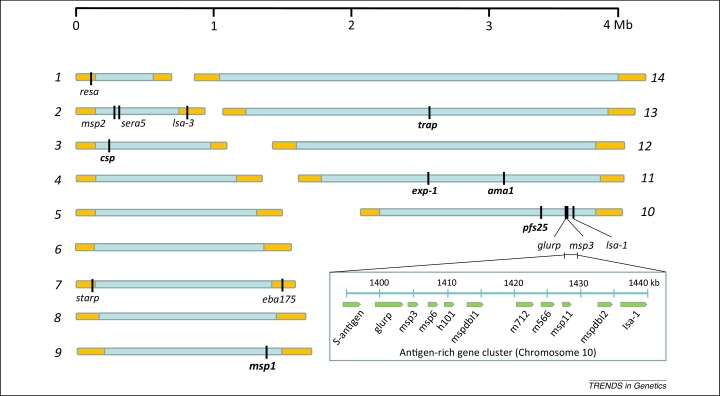
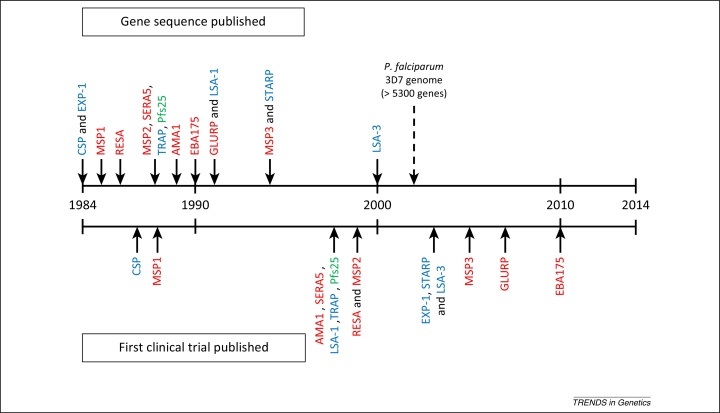
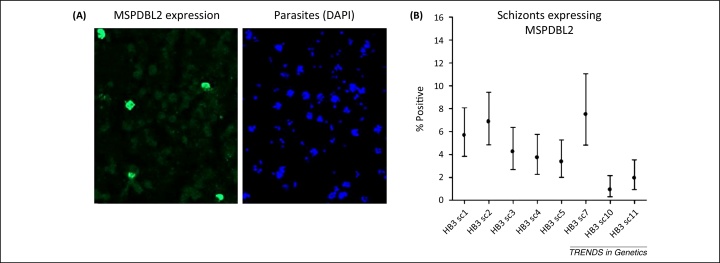
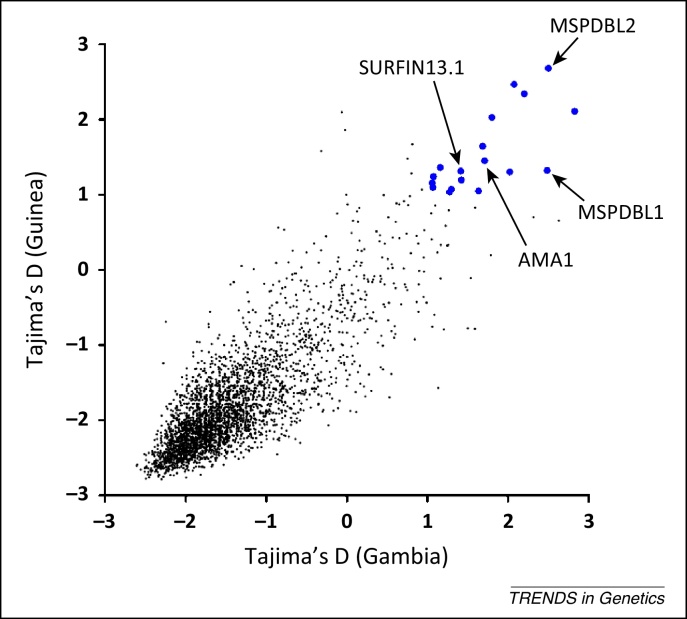
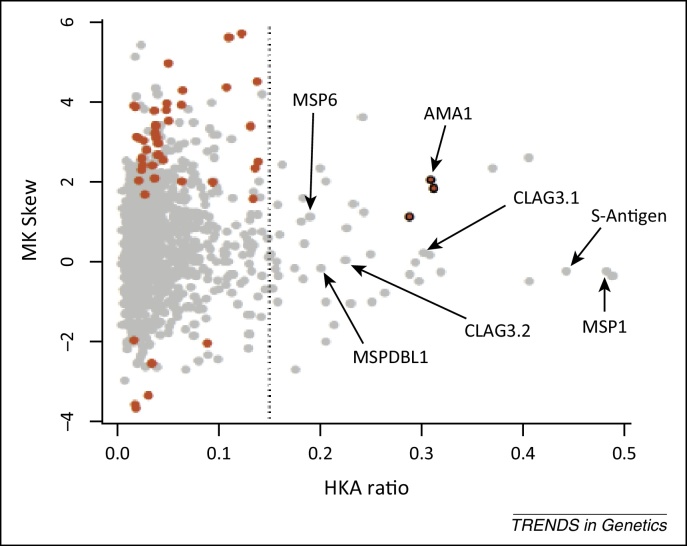
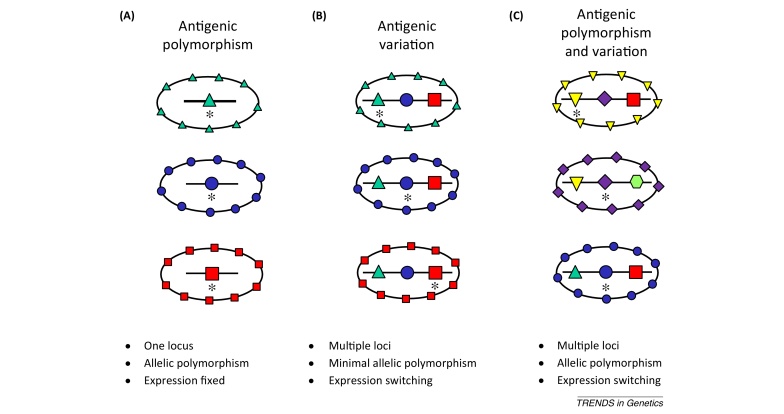
References
-
- Hoffman S.L. From genomics to vaccines: malaria as a model system. Nat. Med. 1998;4:1351–1353. - PubMed
-
- White N.J. Malaria. Lancet. 2014;383:723–735. - PubMed
-
- Birkett A.J. Malaria vaccine R&D in the Decade of Vaccines: breakthroughs, challenges and opportunities. Vaccine. 2013;31(Suppl. 2):B233–B243. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
