

Naturally occurring differences in CENH3 affect chromosome segregation in zygotic mitosis of hybrids
- PMID: 25622028
- PMCID: PMC4314295
- DOI: 10.1371/journal.pgen.1004970
Naturally occurring differences in CENH3 affect chromosome segregation in zygotic mitosis of hybrids
Abstract
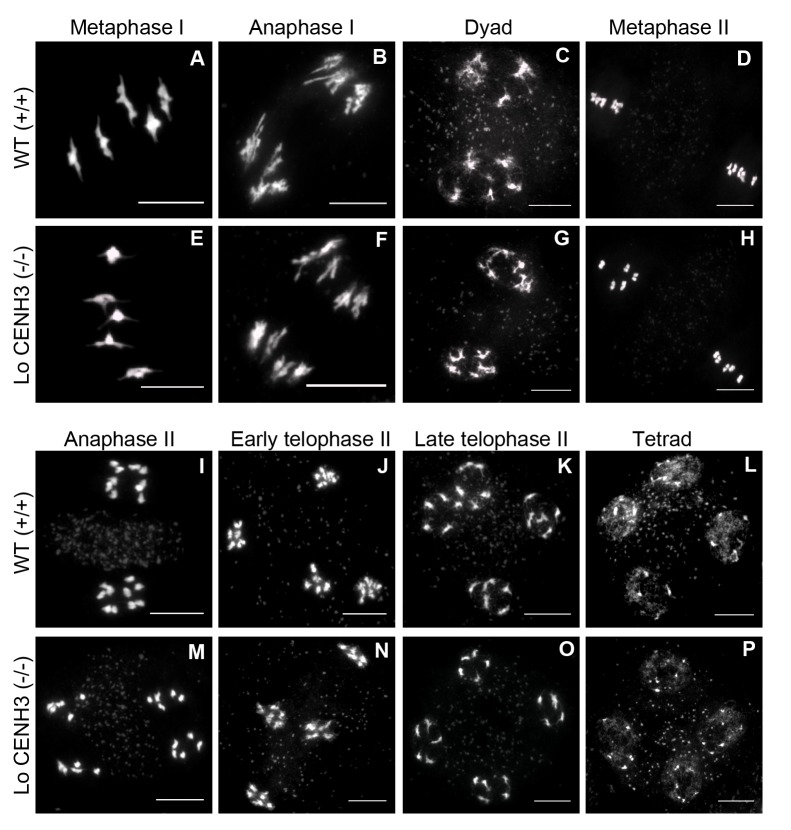
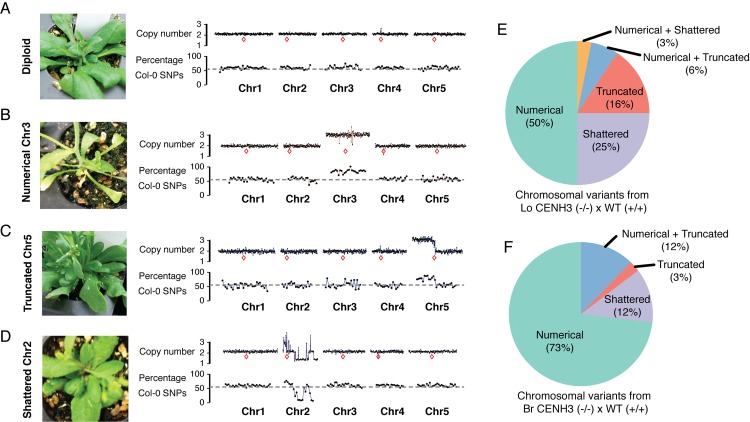
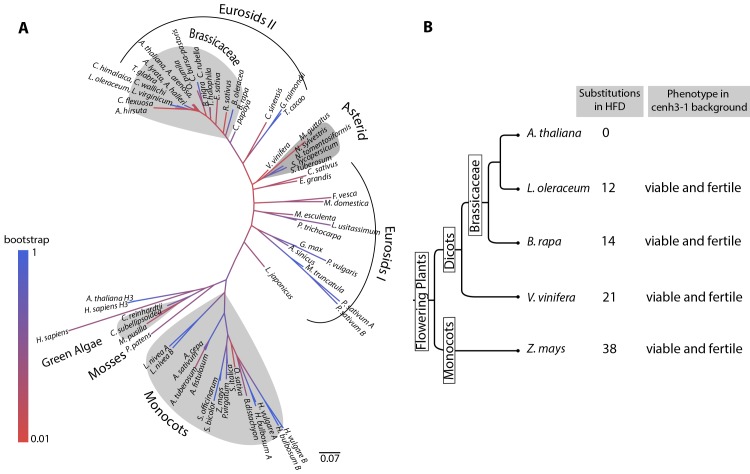
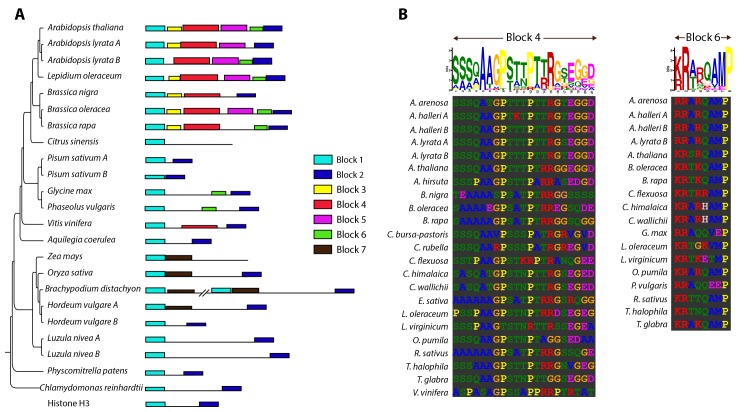
The point of attachment of spindle microtubules to metaphase chromosomes is known as the centromere. Plant and animal centromeres are epigenetically specified by a centromere-specific variant of Histone H3, CENH3 (a.k.a. CENP-A). Unlike canonical histones that are invariant, CENH3 proteins are accumulating substitutions at an accelerated rate. This diversification of CENH3 is a conundrum since its role as the key determinant of centromere identity remains a constant across species. Here, we ask whether naturally occurring divergence in CENH3 has functional consequences. We performed functional complementation assays on cenh3-1, a null mutation in Arabidopsis thaliana, using untagged CENH3s from increasingly distant relatives. Contrary to previous results using GFP-tagged CENH3, we find that the essential functions of CENH3 are conserved across a broad evolutionary landscape. CENH3 from a species as distant as the monocot Zea mays can functionally replace A. thaliana CENH3. Plants expressing variant CENH3s that are fertile when selfed show dramatic segregation errors when crossed to a wild-type individual. The progeny of this cross include hybrid diploids, aneuploids with novel genetic rearrangements and haploids that inherit only the genome of the wild-type parent. Importantly, it is always chromosomes from the plant expressing the divergent CENH3 that missegregate. Using chimeras, we show that it is divergence in the fast-evolving N-terminal tail of CENH3 that is causing segregation errors and genome elimination. Furthermore, we analyzed N-terminal tail sequences from plant CENH3s and discovered a modular pattern of sequence conservation. From this we hypothesize that while the essential functions of CENH3 are largely conserved, the N-terminal tail is evolving to adapt to lineage-specific centromeric constraints. Our results demonstrate that this lineage-specific evolution of CENH3 causes inviability and sterility of progeny in crosses, at the same time producing karyotypic variation. Thus, CENH3 evolution can contribute to postzygotic reproductive barriers.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- du Sart D, Cancilla MR, Earle E, Mao JI, Saffery R, et al. (1997) A functional neo-centromere formed through activation of a latent human centromere and consisting of non-alpha-satellite DNA. Nat Genet 16: 144–153. - PubMed
Publication types
MeSH terms
Substances
Associated data
- BioProject/PRJNA261254
- Actions
- SRA/SRP048570
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
