

Structure-function analysis of VapB4 antitoxin identifies critical features of a minimal VapC4 toxin-binding module
- PMID: 25622615
- PMCID: PMC4352670
- DOI: 10.1128/JB.02508-14
Structure-function analysis of VapB4 antitoxin identifies critical features of a minimal VapC4 toxin-binding module
Abstract
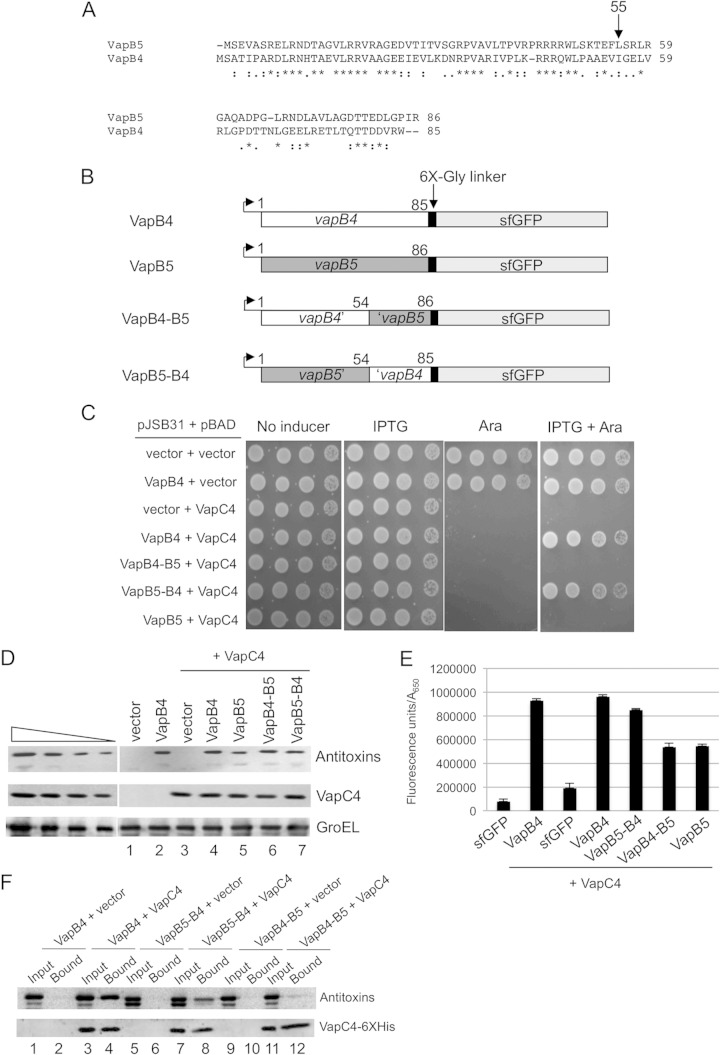
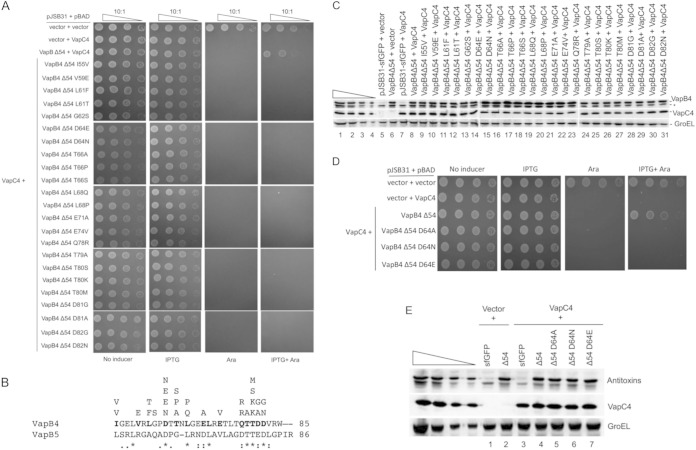
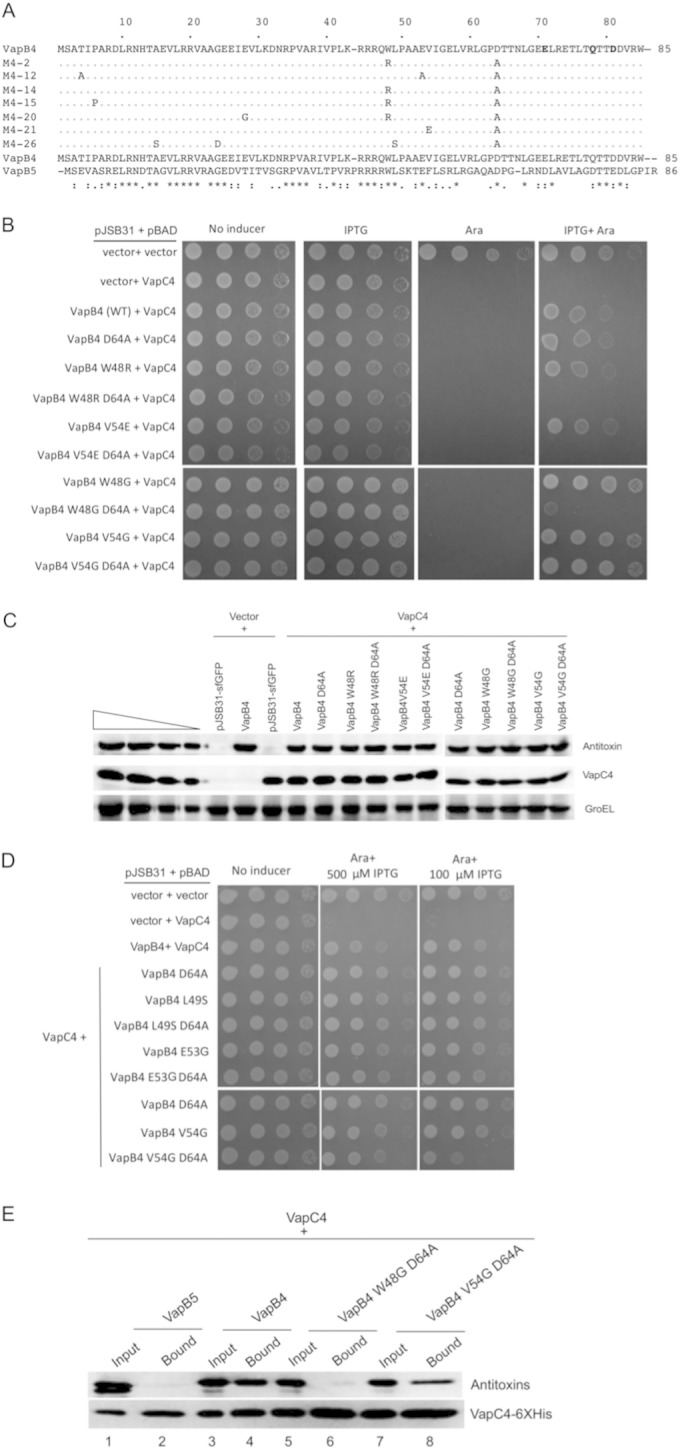
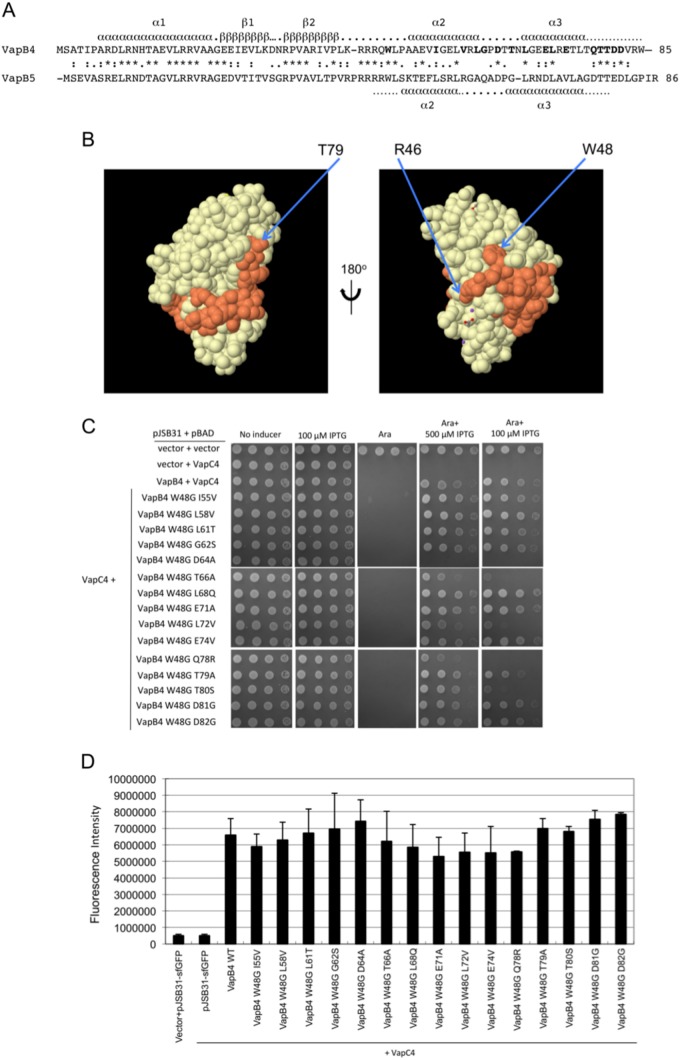
Bacterial toxin-antitoxin systems play a critical role in the regulation of gene expression, leading to developmental changes, reversible dormancy, and cell death. Type II toxin-antitoxin pairs, composed of protein toxins and antitoxins, exist in nearly all bacteria and are classified into six groups on the basis of the structure of the toxins. The VapBC group comprises the most common type II system and, like other toxin-antitoxin systems, functions to elicit dormancy by inhibiting protein synthesis. Activation of toxin function requires protease degradation of the VapB antitoxin, which frees the VapC toxin from the VapBC complex, allowing it to hydrolyze the RNAs required for translation. Generally, type II antitoxins bind with high specificity to their cognate toxins via a toxin-binding domain and endow the complex with DNA-binding specificity via a DNA-binding domain. Despite the ubiquity of VapBC systems and their critical role in the regulation of gene expression, few functional studies have addressed the details of VapB-VapC interactions. Here we report on the results of experiments designed to identify molecular determinants of the specificity of the Mycobacterium tuberculosis VapB4 antitoxin for its cognate VapC4 toxin. The results identify the minimal domain of VapB4 required for this interaction as well as the amino acid side chains required for binding to VapC4. These findings have important implications for the evolution of VapBC toxin-antitoxin systems and their potential as targets of small-molecule protein-protein interaction inhibitors.
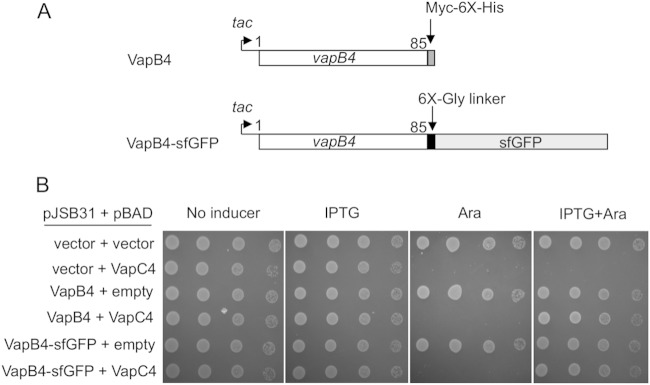
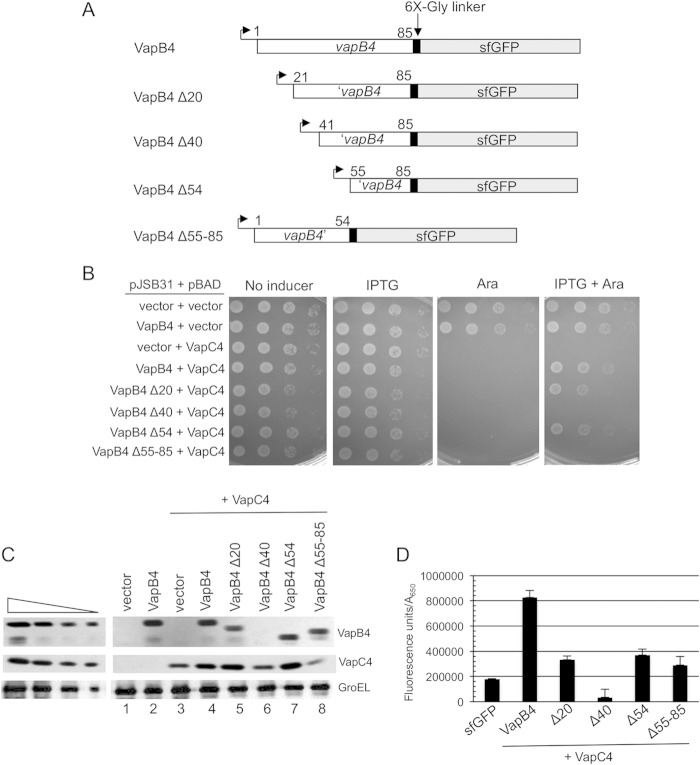
Importance: VapBC toxin-antitoxin pairs are the most widespread type II toxin-antitoxin systems in bacteria, where they are thought to play key roles in stress-induced dormancy and the formation of persisters. The VapB antitoxins are critical to these processes because they inhibit the activity of the toxins and provide the DNA-binding specificity that controls the synthesis of both proteins. Despite the importance of VapB antitoxins and the existence of several VapBC crystal structures, little is known about their functional features in vivo. Here we report the findings of the first comprehensive structure-function analysis of a VapB toxin. The results identify the minimal toxin-binding domain, its modular antitoxin function, and the specific amino acid side chains required for its activity.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
References
-
- Gerdes K, Molin S. 1986. Partitioning of plasmid R1. Structural and functional analysis of the parA locus. J Mol Biol 190:269–279. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
