

Heterogeneity for IGF-II production maintained by public goods dynamics in neuroendocrine pancreatic cancer
- PMID: 25624490
- PMCID: PMC4330744
- DOI: 10.1073/pnas.1414653112
Heterogeneity for IGF-II production maintained by public goods dynamics in neuroendocrine pancreatic cancer
Abstract
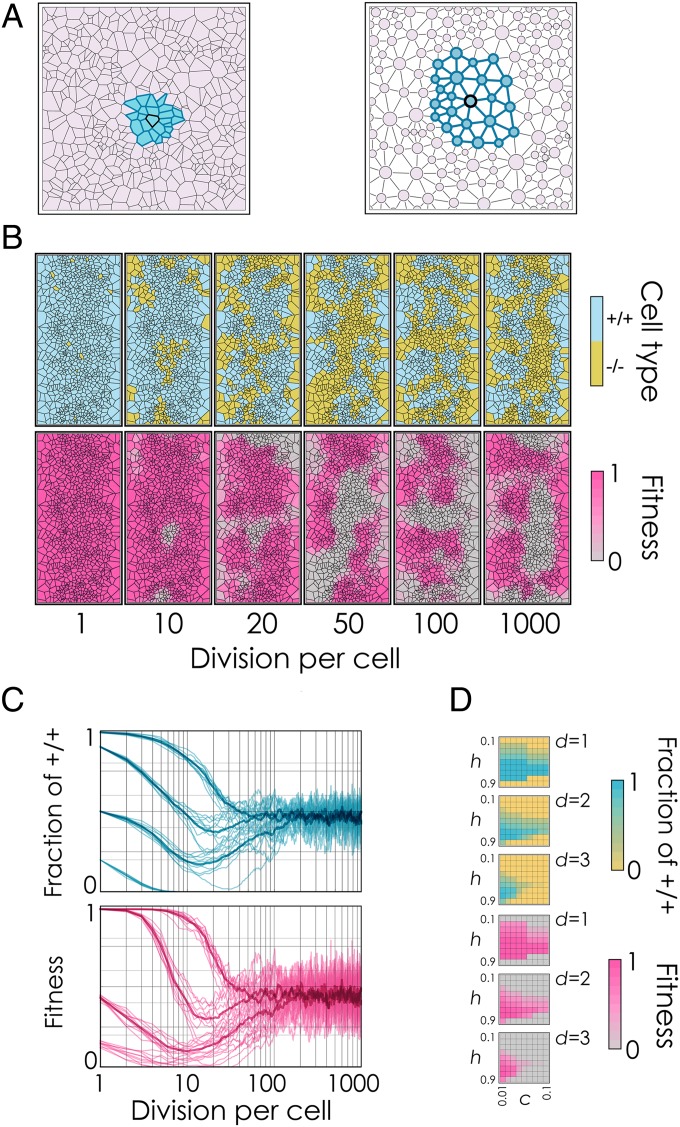
The extensive intratumor heterogeneity revealed by sequencing cancer genomes is an essential determinant of tumor progression, diagnosis, and treatment. What maintains heterogeneity remains an open question because competition within a tumor leads to a strong selection for the fittest subclone. Cancer cells also cooperate by sharing molecules with paracrine effects, such as growth factors, and heterogeneity can be maintained if subclones depend on each other for survival. Without strict interdependence between subclones, however, nonproducer cells can free-ride on the growth factors produced by neighboring producer cells, a collective action problem known in game theory as the "tragedy of the commons," which has been observed in microbial cell populations. Here, we report that similar dynamics occur in cancer cell populations. Neuroendocrine pancreatic cancer (insulinoma) cells that do not produce insulin-like growth factor II (IGF-II) grow slowly in pure cultures but have a proliferation advantage in mixed cultures, where they can use the IGF-II provided by producer cells. We show that, as predicted by evolutionary game theory, producer cells do not go extinct because IGF-II acts as a nonlinear public good, creating negative frequency-dependent selection that leads to a stable coexistence of the two cell types. Intratumor cell heterogeneity can therefore be maintained even without strict interdependence between cell subclones. Reducing the amount of growth factors available within a tumor may lead to a reduction in growth followed by a new equilibrium, which may explain relapse in therapies that target growth factors.
Keywords: evolution; game theory; tumor.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
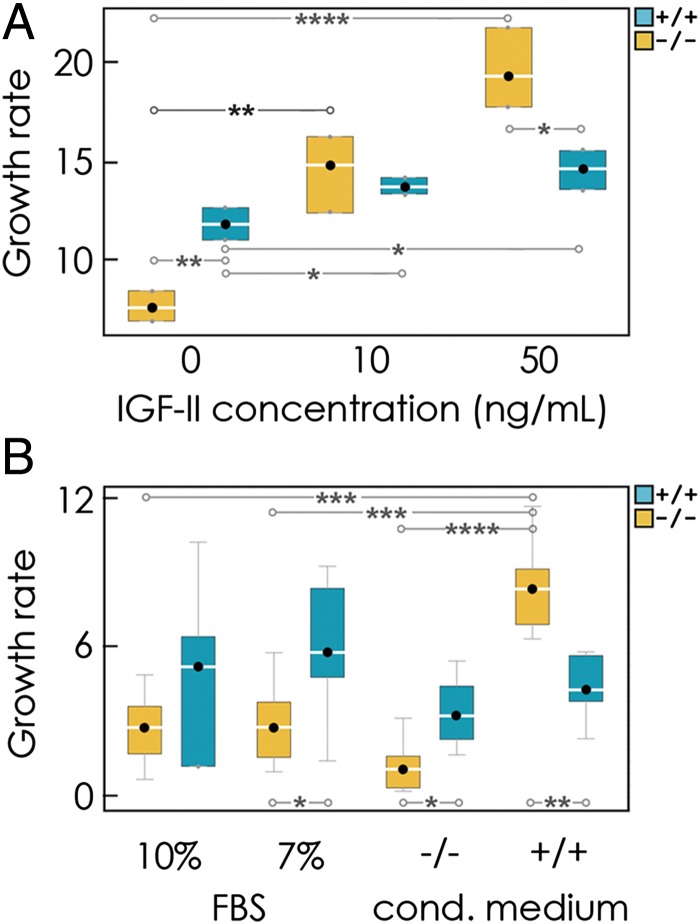
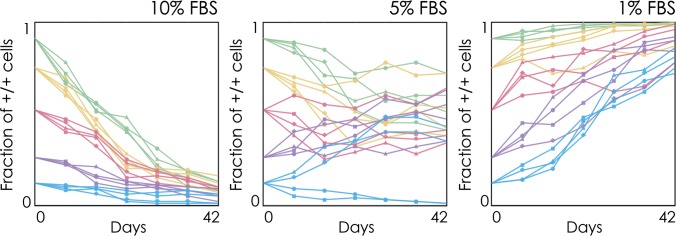
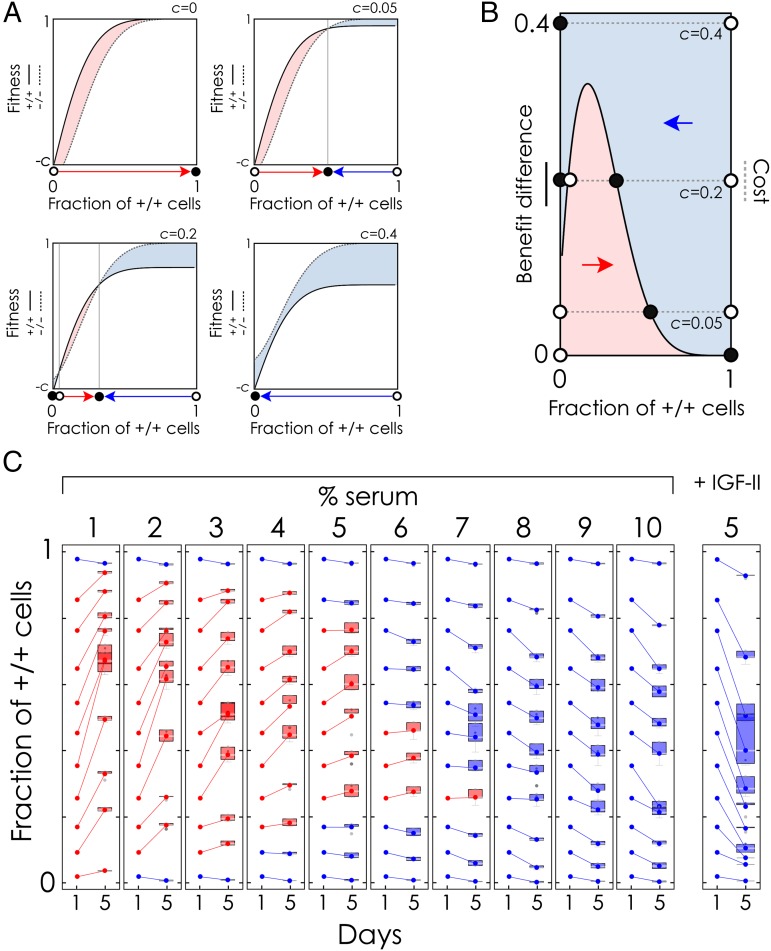
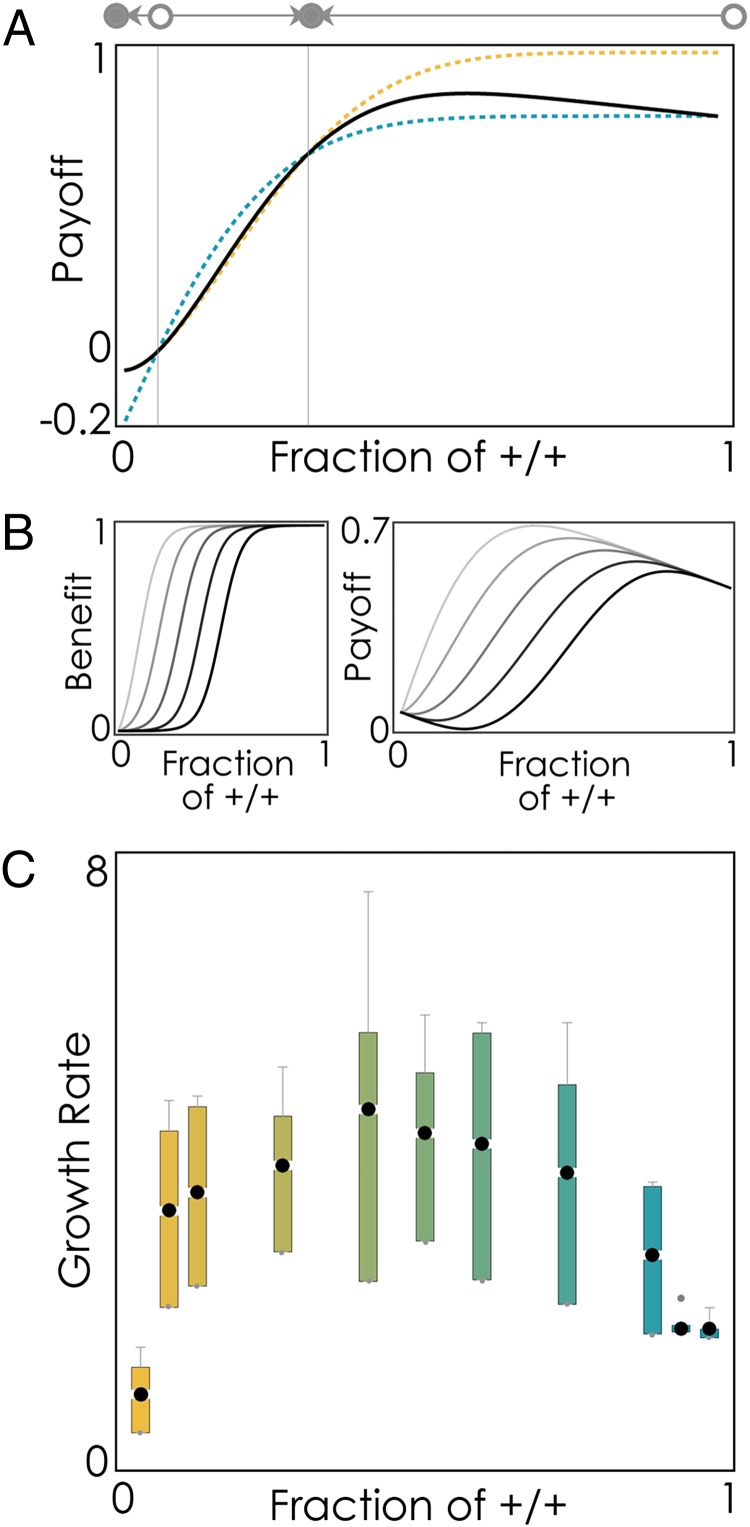
Comment in
-
Reply to Gerlee and Altrock: Diffusion and population size in game theory models of cancer.Proc Natl Acad Sci U S A. 2015 May 26;112(21):E2744. doi: 10.1073/pnas.1505648112. Epub 2015 May 4. Proc Natl Acad Sci U S A. 2015. PMID: 25941413 Free PMC article. No abstract available.
-
Complexity and stability in growing cancer cell populations.Proc Natl Acad Sci U S A. 2015 May 26;112(21):E2742-3. doi: 10.1073/pnas.1505115112. Epub 2015 May 4. Proc Natl Acad Sci U S A. 2015. PMID: 25941414 Free PMC article. No abstract available.
References
-
- Cairns J. Mutation selection and the natural history of cancer. Nature. 1975;255(5505):197–200. - PubMed
-
- Nowell PC. The clonal evolution of tumor cell populations. Science. 1976;194(4260):23–28. - PubMed
-
- Merlo LMF, Pepper JW, Reid BJ, Maley CC. Cancer as an evolutionary and ecological process. Nat Rev Cancer. 2006;6(12):924–935. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
