

RNA methyltransferases involved in 5' cap biosynthesis
- PMID: 25626080
- PMCID: PMC4615557
- DOI: 10.1080/15476286.2015.1004955
RNA methyltransferases involved in 5' cap biosynthesis
Abstract
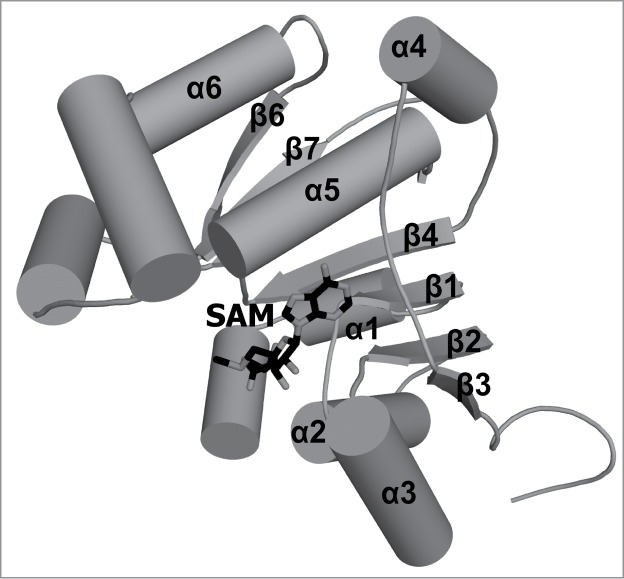
In eukaryotes and viruses that infect them, the 5' end of mRNA molecules, and also many other functionally important RNAs, are modified to form a so-called cap structure that is important for interactions of these RNAs with many nuclear and cytoplasmic proteins. The RNA cap has multiple roles in gene expression, including enhancement of RNA stability, splicing, nucleocytoplasmic transport, and translation initiation. Apart from guanosine addition to the 5' end in the most typical cap structure common to transcripts produced by RNA polymerase II (in particular mRNA), essentially all cap modifications are due to methylation. The complexity of the cap structure and its formation can range from just a single methylation of the unprocessed 5' end of the primary transcript, as in mammalian U6 and 7SK, mouse B2, and plant U3 RNAs, to an elaborate m(7)Gpppm(6,6)AmpAmpCmpm(3)Um structure at the 5' end of processed RNA in trypanosomes, which are formed by as many as 8 methylation reactions. While all enzymes responsible for methylation of the cap structure characterized to date were found to belong to the same evolutionarily related and structurally similar Rossmann Fold Methyltransferase superfamily, that uses the same methyl group donor, S-adenosylmethionine; the enzymes also exhibit interesting differences that are responsible for their distinct functions. This review focuses on the evolutionary classification of enzymes responsible for cap methylation in RNA, with a focus on the sequence relationships and structural similarities and dissimilarities that provide the basis for understanding the mechanism of biosynthesis of different caps in cellular and viral RNAs. Particular attention is paid to the similarities and differences between methyltransferases from human cells and from human pathogens that may be helpful in the development of antiviral and antiparasitic drugs.
Keywords: RNA maturation; RNA modification; antiviral drugs; cap; crystallography; mRNA; methylation; modified nucleotides; post-transcriptional modification; trypanosomes.
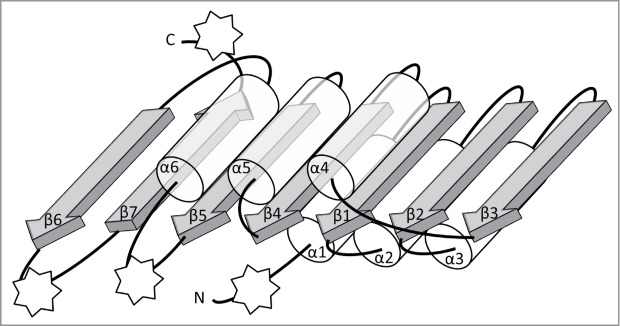
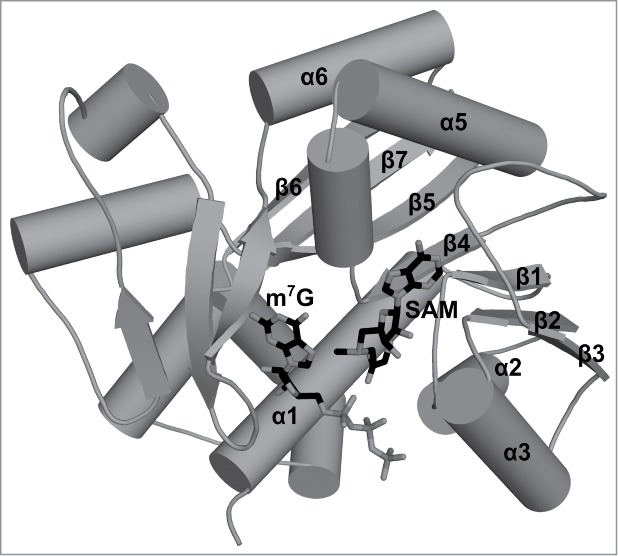
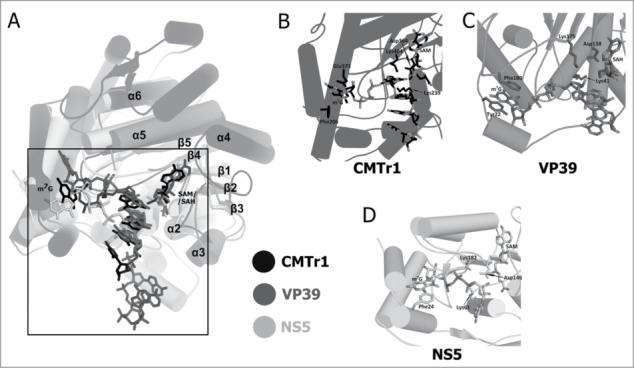
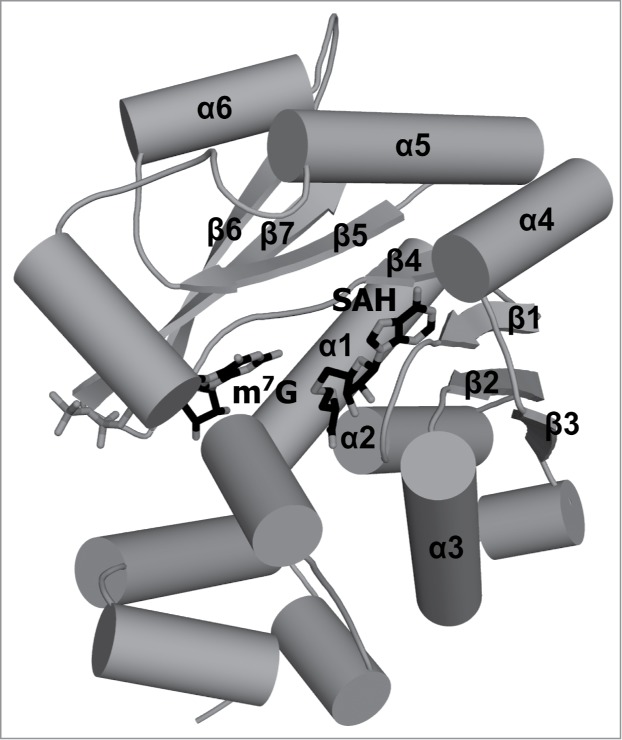
Figures
References
-
- Muthukrishnan S, Filipowicz W, Sierra JM, Both GW, Shatkin AJ, Ochoa S. mRNA methylation and protein synthesis in extracts from embryos of brine shrimp, Artemia salina. J Biol Chem 1975; 250:9336-41; PMID:1194288 - PubMed
-
- Shafer B, Chu C, Shatkin AJ. Human mRNA cap methyltransferase: alternative nuclear localization signal motifs ensure nuclear localization required for viability. Mol Cell Biol 2005; 25:2644-9; PMID:15767670; http://dx.doi.org/10.1128/MCB.25.7.2644-2649.2005 - DOI - PMC - PubMed
-
- Topisirovic I, Svitkin YV, Sonenberg N, Shatkin AJ. Cap and cap-binding proteins in the control of gene expression. Wiley Interdiscip Rev RNA 2011; 2:277-98; PMID:21957010; http://dx.doi.org/10.1002/wrna.52 - DOI - PubMed
-
- Hocine S, Singer RH, Grunwald D. RNA processing and export. Cold Spring Harb Perspect Biol 2010; 2:a000752; PMID:20961978; http://dx.doi.org/10.1101/cshperspect.a000752 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous