

Increased muscle blood supply and transendothelial nutrient and insulin transport induced by food intake and exercise: effect of obesity and ageing
- PMID: 25627798
- PMCID: PMC4933113
- DOI: 10.1113/jphysiol.2014.284513
Increased muscle blood supply and transendothelial nutrient and insulin transport induced by food intake and exercise: effect of obesity and ageing
Abstract
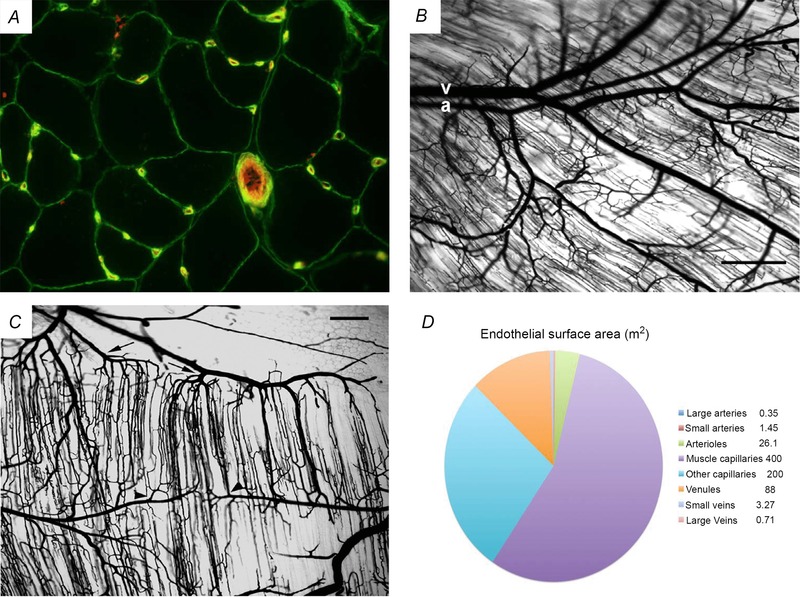
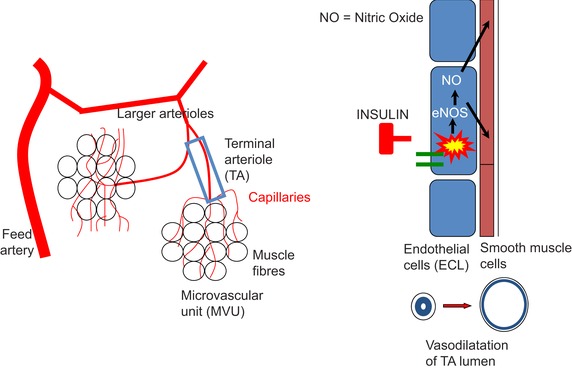
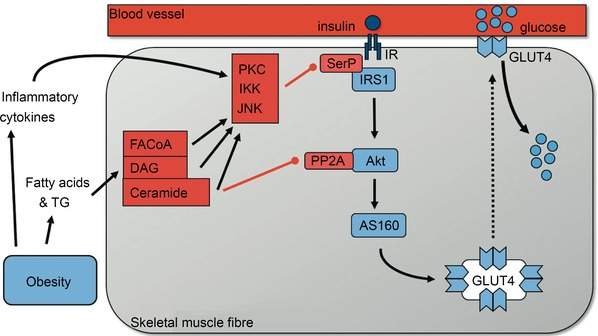
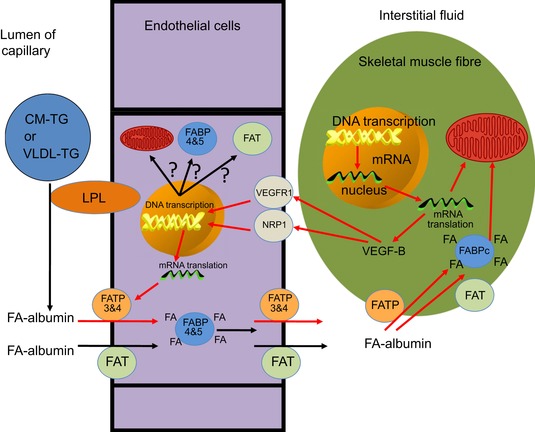
This review concludes that a sedentary lifestyle, obesity and ageing impair the vasodilator response of the muscle microvasculature to insulin, exercise and VEGF-A and reduce microvascular density. Both impairments contribute to the development of insulin resistance, obesity and chronic age-related diseases. A physically active lifestyle keeps both the vasodilator response and microvascular density high. Intravital microscopy has shown that microvascular units (MVUs) are the smallest functional elements to adjust blood flow in response to physiological signals and metabolic demands on muscle fibres. The luminal diameter of a common terminal arteriole (TA) controls blood flow through up to 20 capillaries belonging to a single MVU. Increases in plasma insulin and exercise/muscle contraction lead to recruitment of additional MVUs. Insulin also increases arteriolar vasomotion. Both mechanisms increase the endothelial surface area and therefore transendothelial transport of glucose, fatty acids (FAs) and insulin by specific transporters, present in high concentrations in the capillary endothelium. Future studies should quantify transporter concentration differences between healthy and at risk populations as they may limit nutrient supply and oxidation in muscle and impair glucose and lipid homeostasis. An important recent discovery is that VEGF-B produced by skeletal muscle controls the expression of FA transporter proteins in the capillary endothelium and thus links endothelial FA uptake to the oxidative capacity of skeletal muscle, potentially preventing lipotoxic FA accumulation, the dominant cause of insulin resistance in muscle fibres.
© 2015 The Authors. The Journal of Physiology © 2015 The Physiological Society.
Figures
Comment in
-
Impact of physical activity, ageing, obesity and metabolic syndrome on muscle microvascular perfusion and endothelial metabolism.J Physiol. 2016 Apr 15;594(8):2205-6. doi: 10.1113/JP270525. J Physiol. 2016. PMID: 27079629 Free PMC article. No abstract available.
References
-
- Bansal S, Buring JE, Rifai N, Mora S, Sacks FM & Ridker PM (2007). Fasting compared with nonfasting triglycerides and risk of cardiovascular events in women. JAMA 298, 309–316. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous