

Neurogenesis and growth factors expression after complete spinal cord transection in Pleurodeles waltlii
- PMID: 25628538
- PMCID: PMC4292736
- DOI: 10.3389/fncel.2014.00458
Neurogenesis and growth factors expression after complete spinal cord transection in Pleurodeles waltlii
Abstract
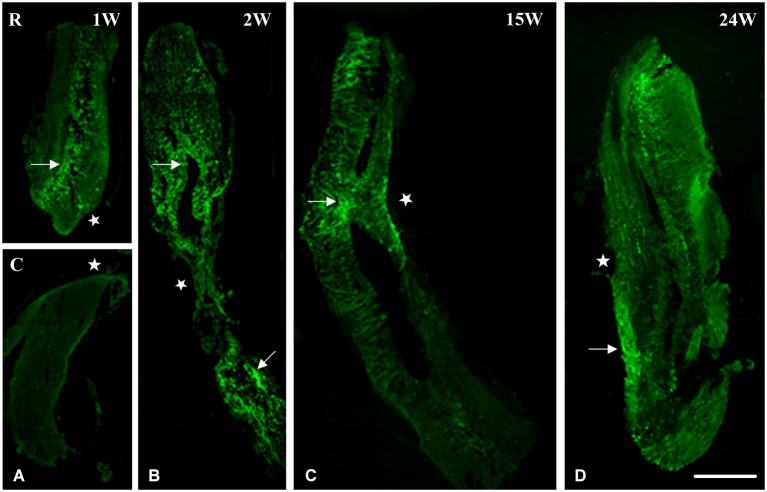
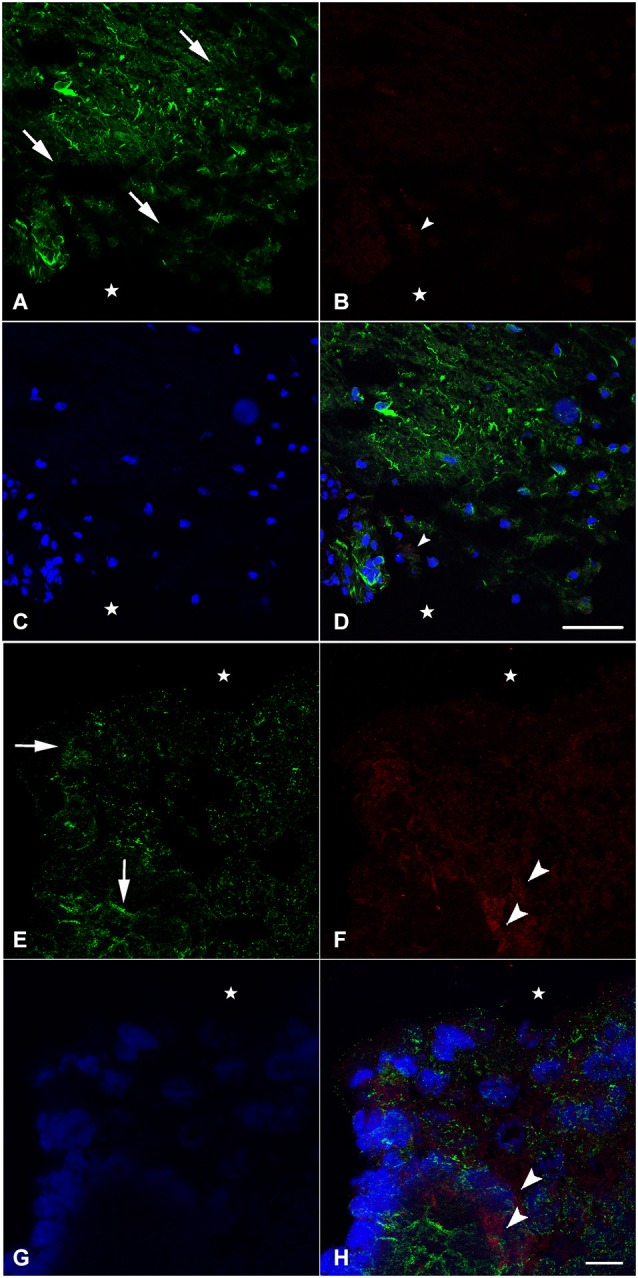
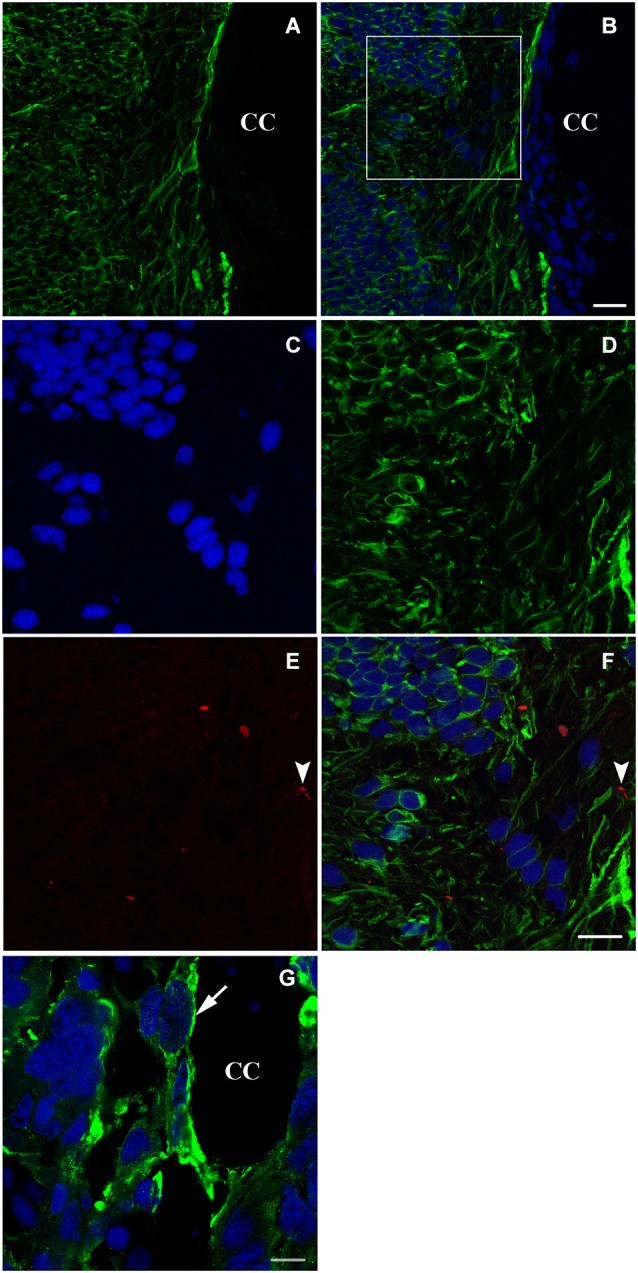
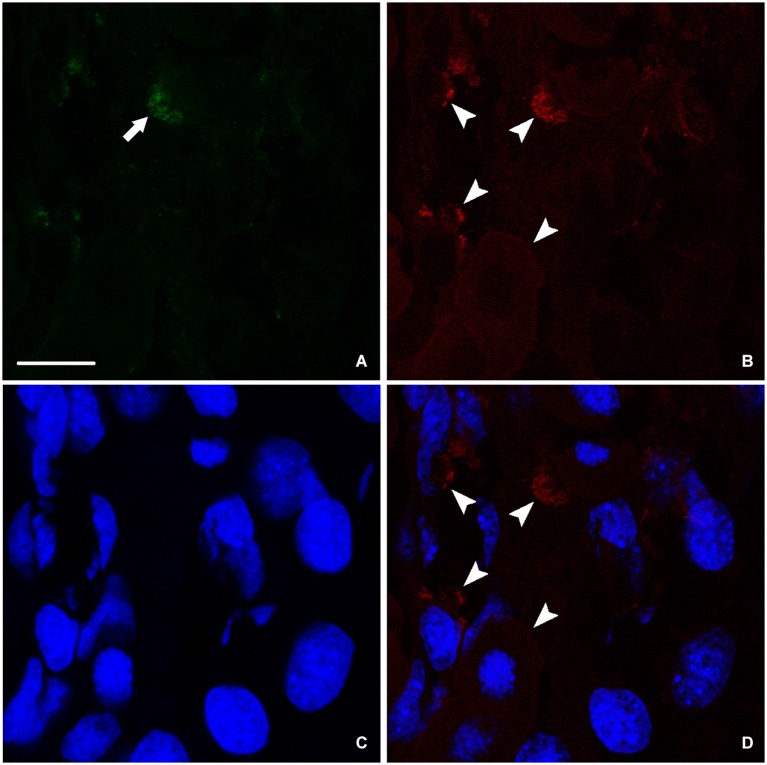
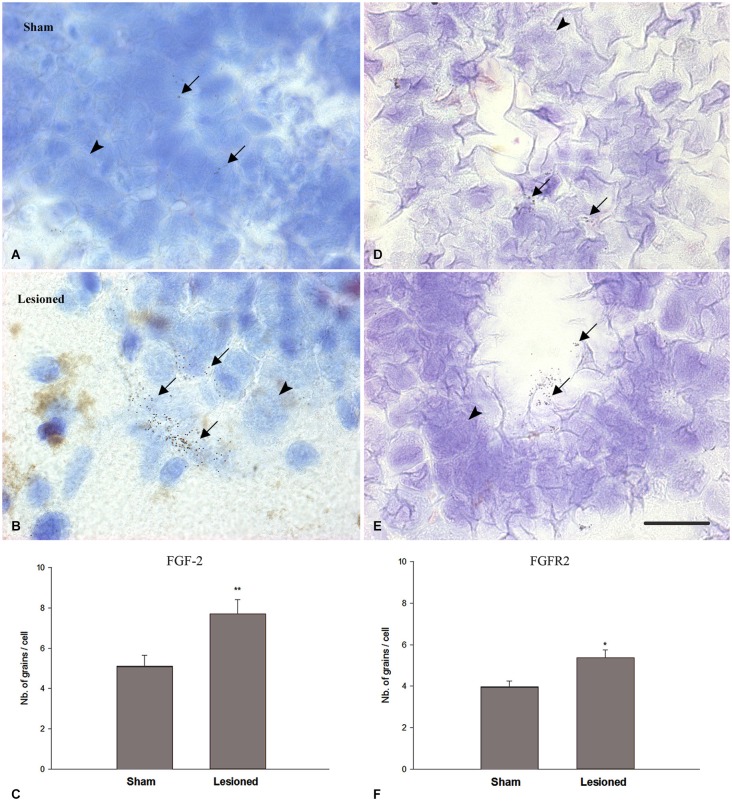
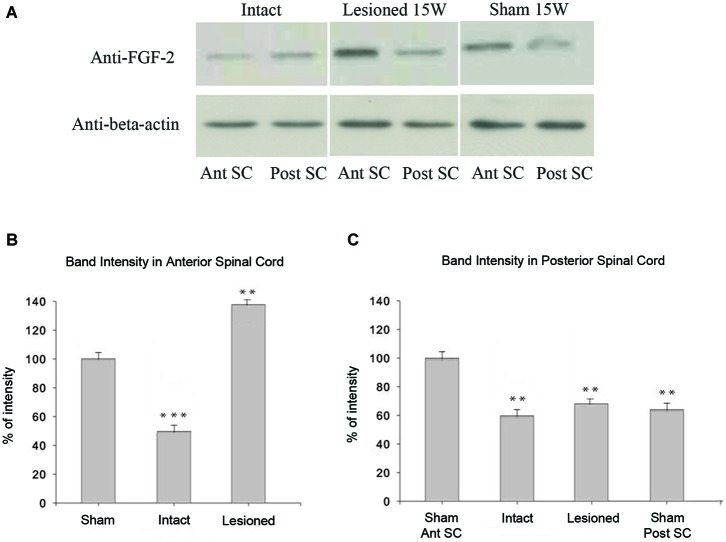
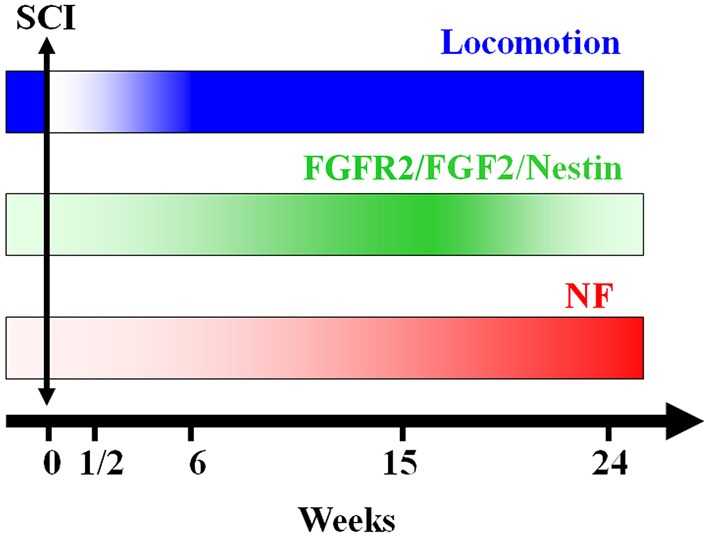
Following spinal lesion, connections between the supra-spinal centers and spinal neuronal networks can be disturbed, which causes the deterioration or even the complete absence of sublesional locomotor activity. In mammals, possibilities of locomotion restoration are much reduced since descending tracts either have very poor regenerative ability or do not regenerate at all. However, in lower vertebrates, there is spontaneous locomotion recuperation after complete spinal cord transection at the mid-trunk level. This phenomenon depends on a translesional descending axon re-growth originating from the brainstem. On the other hand, cellular and molecular mechanisms underlying spinal cord regeneration and in parallel, locomotion restoration of the animal, are not well known. Fibroblast growth factor 2 (FGF-2) plays an important role in different processes such as neural induction, neuronal progenitor proliferation and their differentiation. Studies had shown an over expression of this growth factor after tail amputation. Nestin, a protein specific for intermediate filaments, is considered an early marker for neuronal precursors. It has been recently shown that its expression increases after tail transection in urodeles. Using this marker and western blots, our results show that the number of FGF-2 and FGFR2 mRNAs increases and is correlated with an increase in neurogenesis especially in the central canal lining cells immediately after lesion. This study also confirms that spinal cord re-growth through the lesion site initially follows a rostrocaudal direction. In addition to its role known in neuronal differentiation, FGF-2 could be implicated in the differentiation of ependymal cells into neuronal progenitors.
Keywords: gap replacement; growth factors; locomotion recovery; neurogenesis; spinal cord.
Figures
Similar articles
-
Fibroblast growth factor-2 mRNA expression in the brainstem and spinal cord of normal and chronic spinally transected urodeles.J Neurosci Res. 2008 Nov 15;86(15):3348-58. doi: 10.1002/jnr.21776. J Neurosci Res. 2008. PMID: 18627027
-
FGF-2 Up-regulation and proliferation of neural progenitors in the regenerating amphibian spinal cord in vivo.Dev Biol. 2000 Sep 15;225(2):381-91. doi: 10.1006/dbio.2000.9843. Dev Biol. 2000. PMID: 10985857
-
Differential regulation of fibroblast growth factor receptors in the regenerating amphibian spinal cord in vivo.Neuroscience. 2002;114(4):837-48. doi: 10.1016/s0306-4522(02)00321-4. Neuroscience. 2002. PMID: 12379240
-
Transplants and neurotrophic factors increase regeneration and recovery of function after spinal cord injury.Prog Brain Res. 2002;137:257-73. doi: 10.1016/s0079-6123(02)37020-1. Prog Brain Res. 2002. PMID: 12440372 Review.
-
Progenitors in the Ependyma of the Spinal Cord: A Potential Resource for Self-Repair After Injury.Adv Exp Med Biol. 2017;1015:241-264. doi: 10.1007/978-3-319-62817-2_13. Adv Exp Med Biol. 2017. PMID: 29080030 Review.
References
-
- Alibardi L., Meyer-Rochow V. B. (1988). Ultrastructure of the neural component of the regenerating spinal cord in the tails of three species of New Zealand lizards. N. Z. J. Zool. 15, 535–550 10.1080/03014223.1988.10422633 - DOI
-
- Anderson M. J., Choy C. Y., Waxman S. G. (1986). Self-organization of ependyma in regenerating teleost spinal cord: evidence from serial section reconstructions. J. Embryol. Exp. Morphol. 96, 1–18. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous