

Spatiotemporal characteristics and pharmacological modulation of multiple gamma oscillations in the CA1 region of the hippocampus
- PMID: 25628540
- PMCID: PMC4290596
- DOI: 10.3389/fncir.2014.00150
Spatiotemporal characteristics and pharmacological modulation of multiple gamma oscillations in the CA1 region of the hippocampus
Abstract
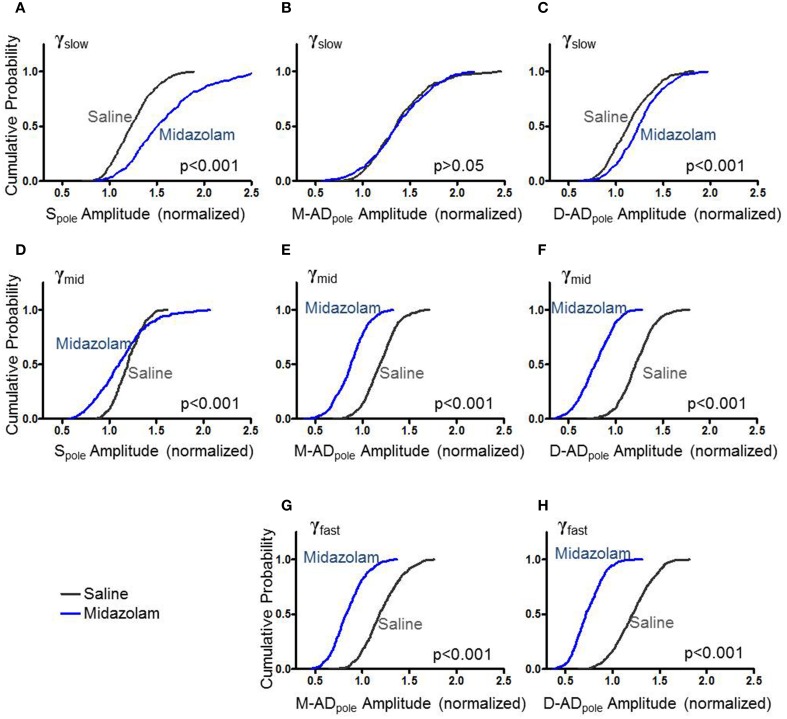
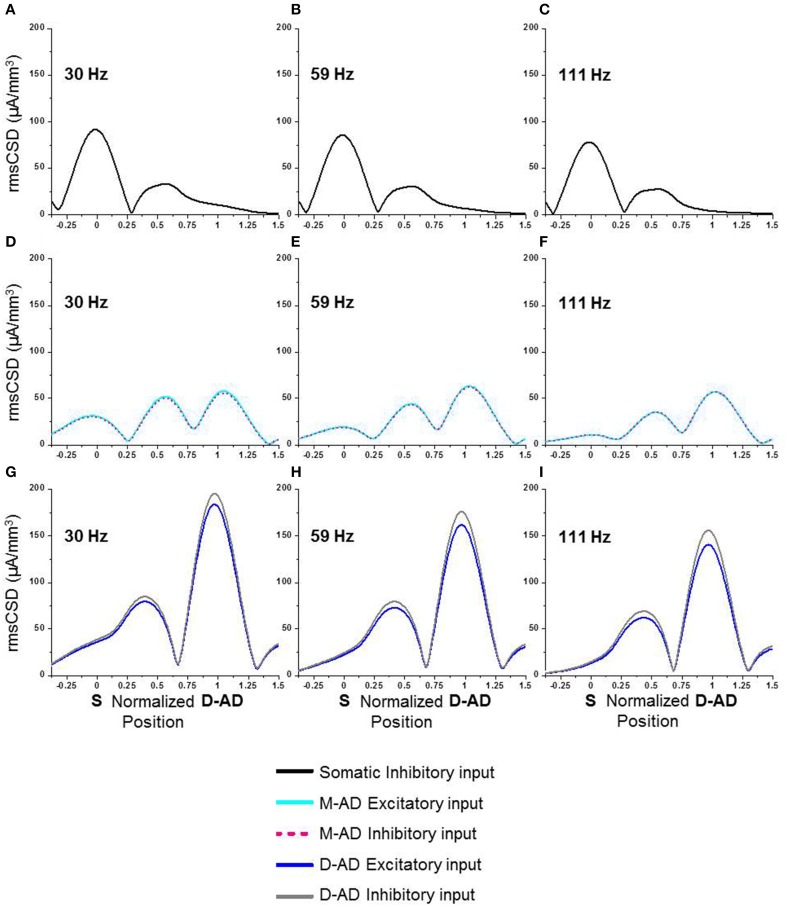
Multiple components of "γ-oscillations" between 30-170 Hz in the CA1 region of the hippocampus have been described, based on their coherence with oscillations in other brain regions and on their cross-frequency coupling with local θ-oscillations. However, it remains unclear whether the different sub-bands are generated by a single broadband oscillator coupled to multiple external inputs, or by separate oscillators that incorporate distinct circuit elements. To distinguish between these possibilities, we used high-density linear array recording electrodes in awake behaving mice to examine the spatiotemporal characteristics of γ-oscillations and their responses to midazolam and atropine. We characterized oscillations using current source density (CSD) analysis, and measured θ-γ phase-amplitude coupling by cross frequency coupling (CFC) analysis. Prominent peaks were present in the CSD signal in the mid- and distal apical dendritic layers at all frequencies, and at stratum pyramidale for γ(slow) (30-45 Hz) and γ(mid) (50-90 Hz), but not γ(fast) (90-170 Hz) oscillations. Differences in the strength and timing of θ-γ(slow) and θ-γ(mid) cross frequency coupling, and a lack of coupling at the soma and mid-apical region for γ(fast) oscillations, indicated that separate circuit components generate the three sub-bands. Midazolam altered CSD amplitudes and cross-frequency coupling in a lamina- and frequency specific manner, providing further evidence for separate generator circuits. Atropine altered CSD amplitudes and θ-γ CFC uniformly at all locations. Simulations using a detailed compartmental model were consistent with γ(slow) and γ(mid) oscillations driven primarily by inputs at the mid-apical dendrites, and γ(fast) at the distal apical dendrite. Our results indicate that multiple distinct local circuits generate γ-oscillations in the CA1 region of the hippocampus, and provide detailed information about their spatiotemporal characteristics.
Keywords: atropine; compartmental modeling; cross frequency coupling; current source density analysis; gamma oscillations; midazolam; theta oscillations.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous