

Dorsal root ganglion myeloid zinc finger protein 1 contributes to neuropathic pain after peripheral nerve trauma
- PMID: 25630025
- PMCID: PMC4366285
- DOI: 10.1097/j.pain.0000000000000103
Dorsal root ganglion myeloid zinc finger protein 1 contributes to neuropathic pain after peripheral nerve trauma
Erratum in
-
Dorsal root ganglion myeloid zinc finger protein 1 contributes to neuropathic pain after peripheral nerve trauma: erratum.Pain. 2022 Feb 1;163(2):e384. doi: 10.1097/j.pain.0000000000002563. Pain. 2022. PMID: 35029603 No abstract available.
Abstract
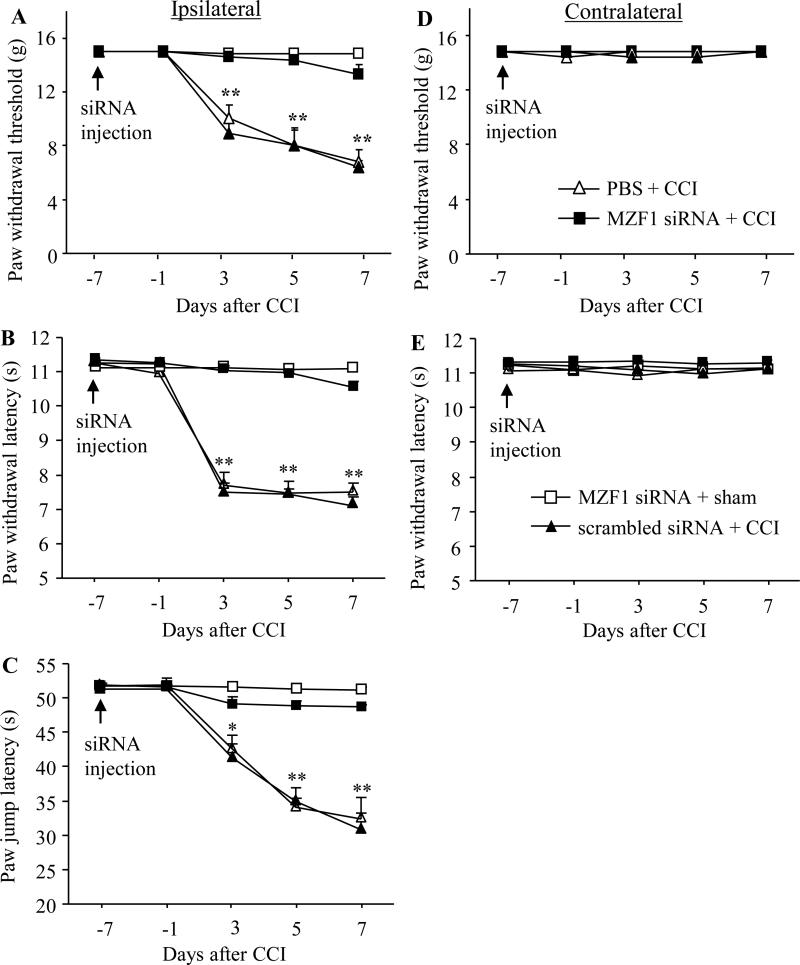
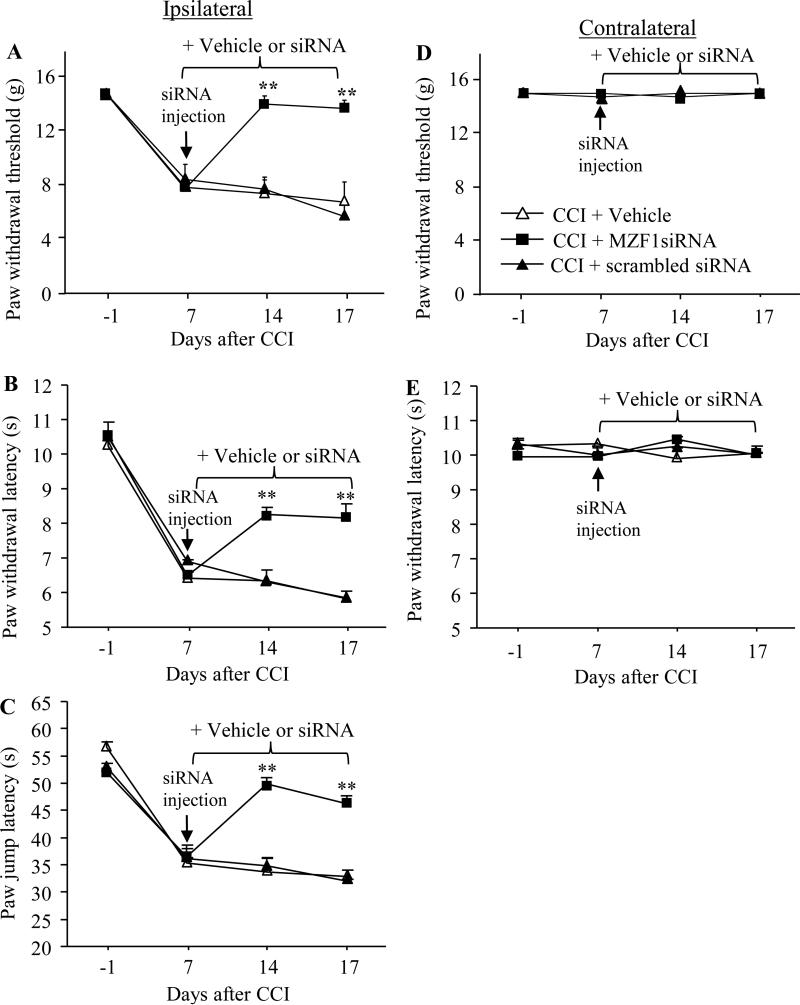
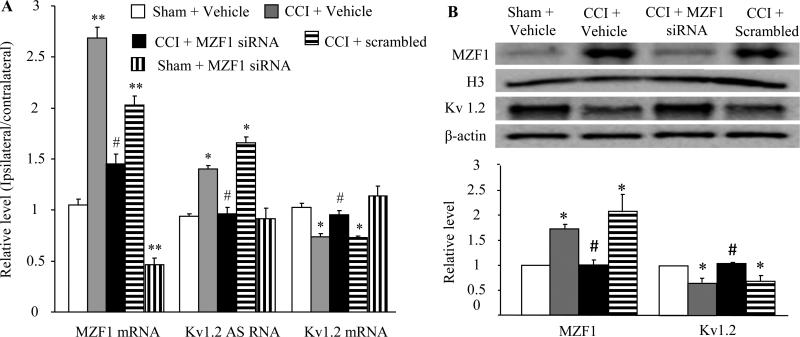
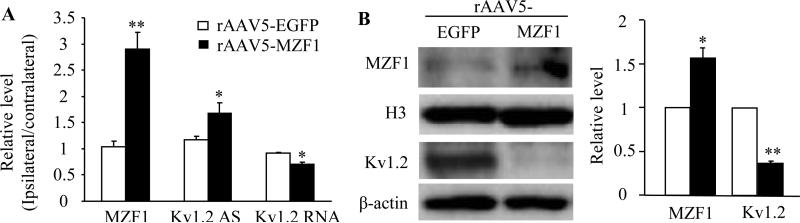
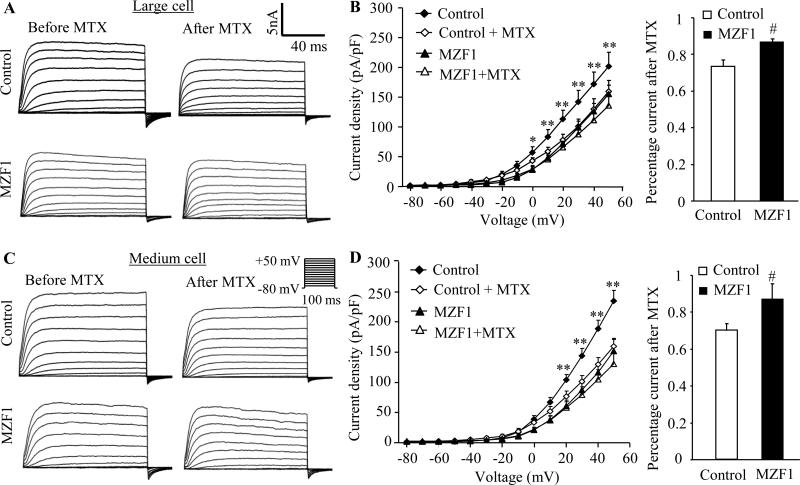
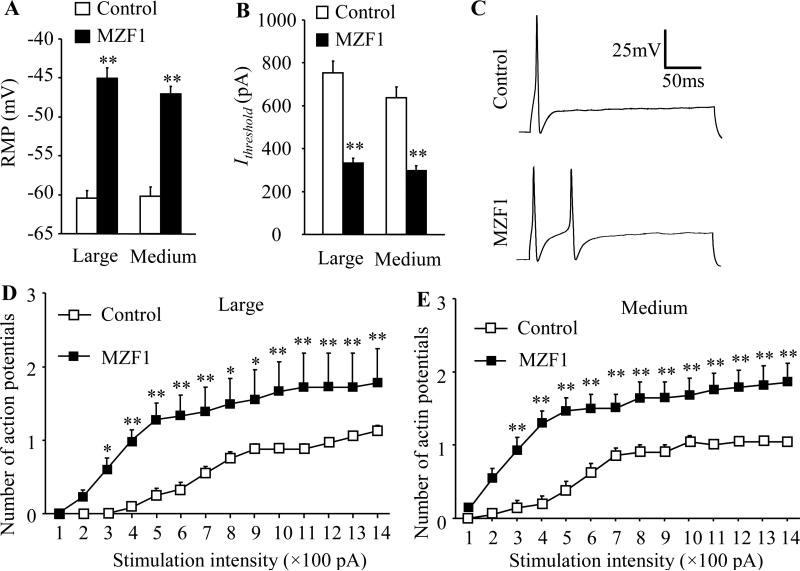
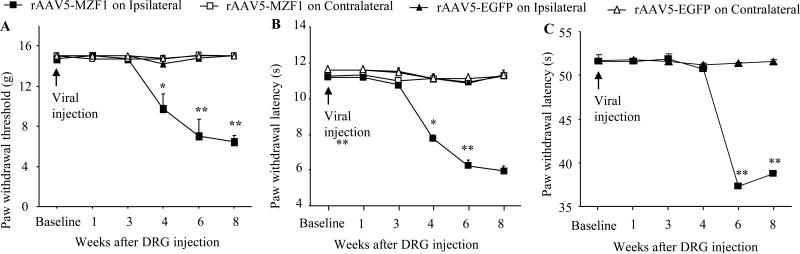
Peripheral nerve injury-induced changes in gene transcription and translation in primary sensory neurons of the dorsal root ganglion (DRG) are considered to contribute to neuropathic pain genesis. Transcription factors control gene expression. Peripheral nerve injury increases the expression of myeloid zinc finger protein 1 (MZF1), a transcription factor, and promotes its binding to the voltage-gated potassium 1.2 (Kv1.2) antisense (AS) RNA gene in the injured DRG. However, whether DRG MZF1 participates in neuropathic pain is still unknown. Here, we report that blocking the nerve injury-induced increase of DRG MZF1 through microinjection of MZF1 siRNA into the injured DRG attenuated the initiation and maintenance of mechanical, cold, and thermal pain hypersensitivities in rats with chronic constriction injury (CCI) of the sciatic nerve, without affecting locomotor functions and basal responses to acute mechanical, heat, and cold stimuli. Mimicking the nerve injury-induced increase of DRG MZF1 through microinjection of recombinant adeno-associated virus 5 expressing full-length MZF1 into the DRG produced significant mechanical, cold, and thermal pain hypersensitivities in naive rats. Mechanistically, MZF1 participated in CCI-induced reductions in Kv1.2 mRNA and protein and total Kv current and the CCI-induced increase in neuronal excitability through MZF1-triggered Kv1.2 AS RNA expression in the injured DRG neurons. MZF1 is likely an endogenous trigger of neuropathic pain and might serve as a potential target for preventing and treating this disorder.
Figures
References
-
- Abrahamsen B, Zhao J, Asante CO, Cendan CM, Marsh S, Martinez-Barbera JP, Nassar MA, Dickenson AH, Wood JN. The cell and molecular basis of mechanical, cold, and inflammatory pain. Science. 2008;321:702–705. - PubMed
-
- Amaya F, Wang H, Costigan M, Allchorne AJ, Hatcher JP, Egerton J, Stean T, Morisset V, Grose D, Gunthorpe MJ, Chessell IP, Tate S, Green PJ, Woolf CJ. The voltage-gated sodium channel Na(v)1.9 is an effector of peripheral inflammatory pain hypersensitivity. J Neurosci. 2006;26:12852–12860. - PMC - PubMed
-
- Bennett GJ, Xie YK. Af peripheral mononeuropathy in rat that produces disorders of pain sensation like those seen in man. Pain. 1988;33:87–107. - PubMed
-
- Castle NA, London DO, Creech C, Fajloun Z, Stocker JW, Sabatier JM. Maurotoxin: a potent inhibitor of intermediate conductance Ca2+-activated potassium channels. Mol Pharmacol. 2003;63:409–418. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
