

Micro- and nano-structural details of a spider's filter for substrate vibrations: relevance for low-frequency signal transmission
- PMID: 25631567
- PMCID: PMC4345480
- DOI: 10.1098/rsif.2014.1111
Micro- and nano-structural details of a spider's filter for substrate vibrations: relevance for low-frequency signal transmission
Abstract
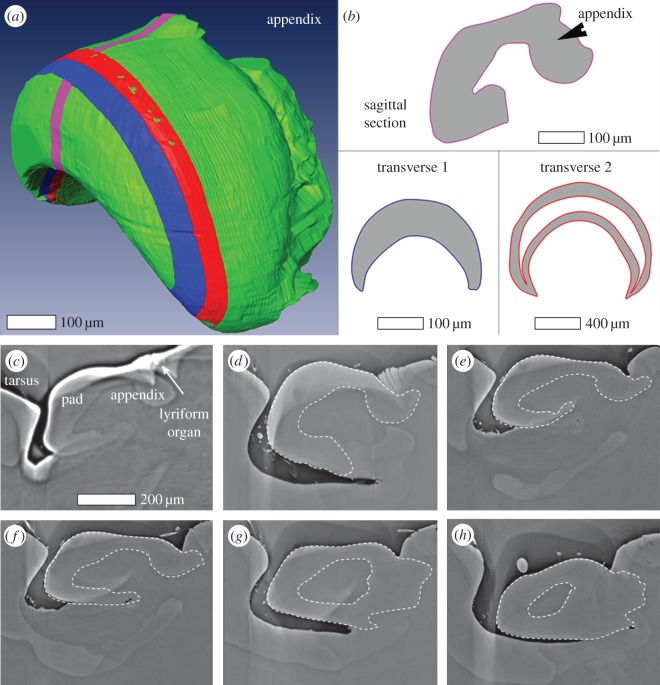
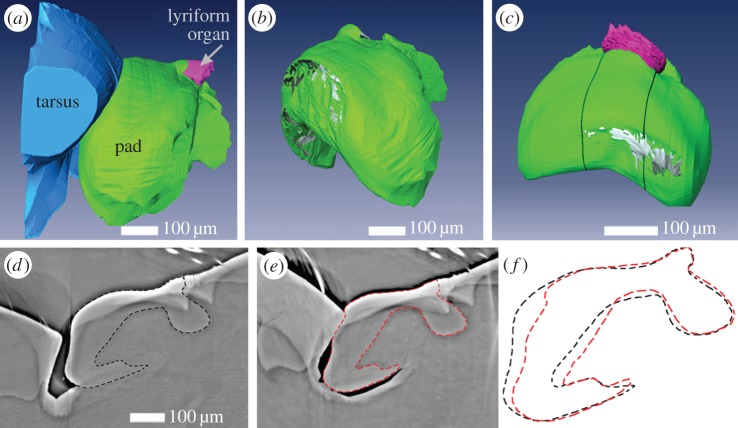
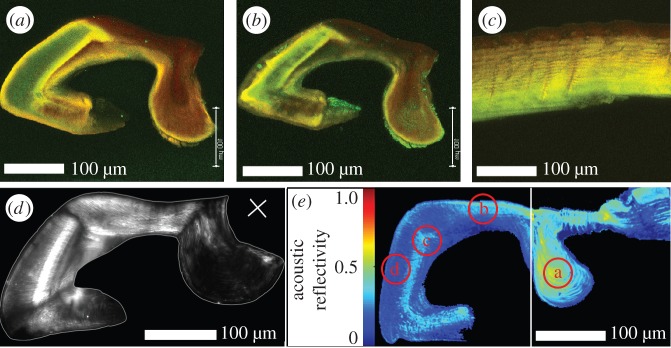
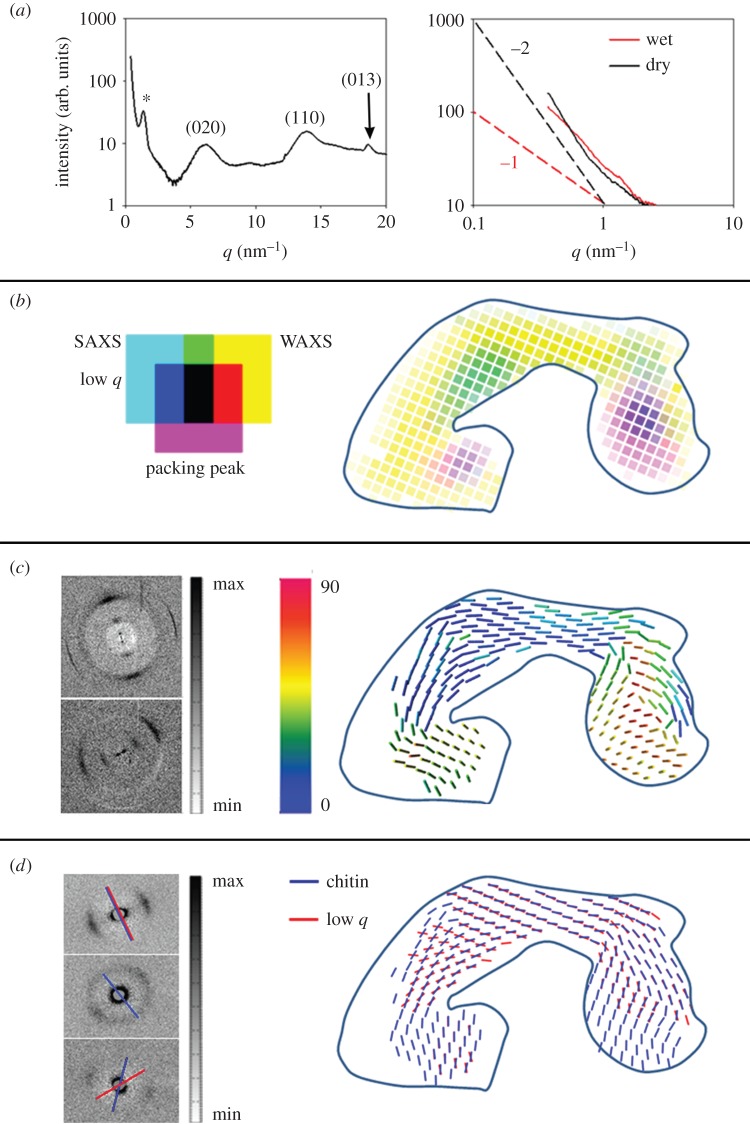
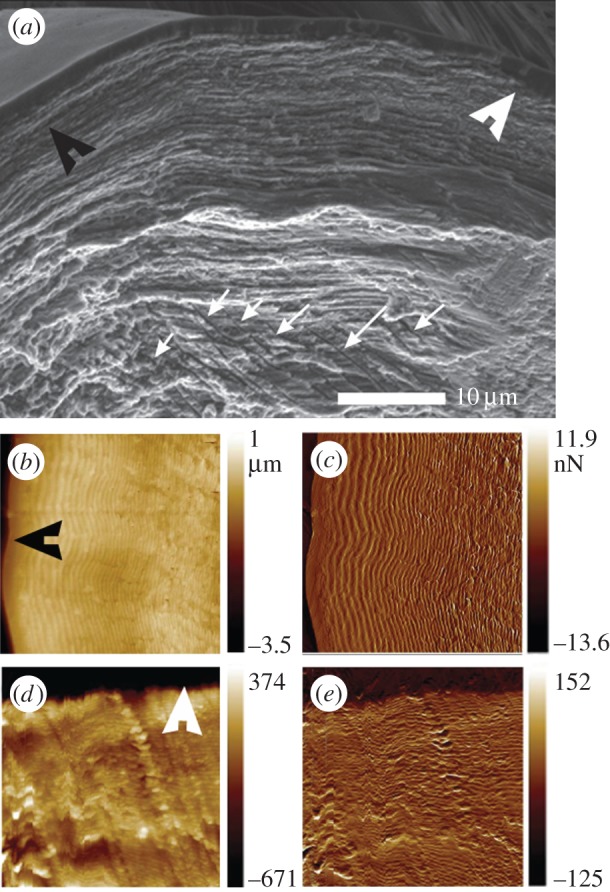
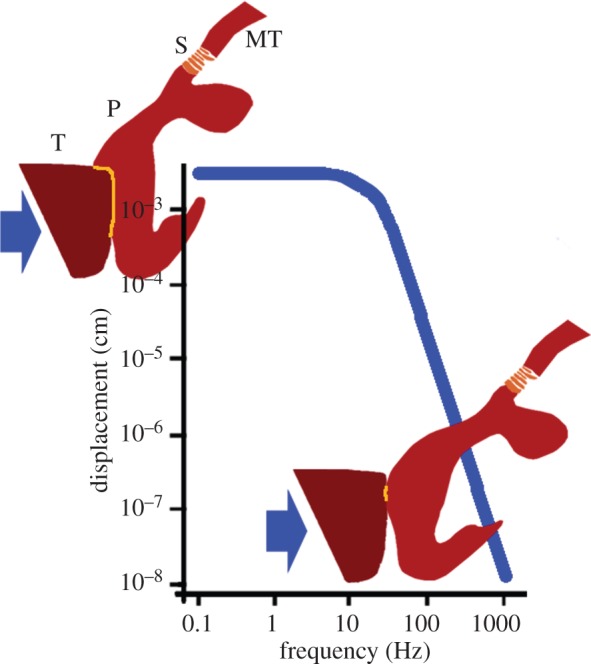
The metatarsal lyriform organ of the Central American wandering spider Cupiennius salei is its most sensitive vibration detector. It is able to sense a wide range of vibration stimuli over four orders of magnitude in frequency between at least as low as 0.1 Hz and several kilohertz. Transmission of the vibrations to the slit organ is controlled by a cuticular pad in front of it. While the mechanism of high-frequency stimulus transfer (above ca 40 Hz) is well understood and related to the viscoelastic properties of the pad's epicuticle, it is not yet clear how low-frequency stimuli (less than 40 Hz) are transmitted. Here, we study how the pad material affects the pad's mechanical properties and thus its role in the transfer of the stimulus, using a variety of experimental techniques, such as X-ray micro-computed tomography for three-dimensional imaging, X-ray scattering for structural analysis, and atomic force microscopy and scanning electron microscopy for surface imaging. The mechanical properties were investigated using scanning acoustic microscopy and nanoindentation. We show that large tarsal deflections cause large deformation in the distal highly hydrated part of the pad. Beyond this region, a sclerotized region serves as a supporting frame which resists the deformation and is displaced to push against the slits, with displacement values considerably scaled down to only a few micrometres. Unravelling the structural arrangement in such specialized structures may provide conceptual ideas for the design of new materials capable of controlling a technical sensor's specificity and selectivity, which is so typical of biological sensors.
Keywords: chitin; mechanosensing; spiders.
Figures

References
-
- Barth FG. 2002. A spider's world: senses and behavior. Berlin, Germany: Springer.
-
- Barth FG. 1972. Die Physiologie der Spaltsinnesorgane. 2. Funktionelle Morphologie eines Mechanorezeptors. J. Comp. Physiol. 81, 159–186. (10.1007/BF00696631) - DOI
-
- Barth FG. 1972. Die Physiologie der Spaltsinnesorgane. 1. Modellversuche zur Rolle des cuticularen Spaltes beim Reiztransport. J. Comp. Physiol. 78, 315–336. (10.1007/BF00697661) - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources