

Size-dependent leak of soluble and membrane proteins through the yeast nuclear pore complex
- PMID: 25631821
- PMCID: PMC4454183
- DOI: 10.1091/mbc.E14-07-1175
Size-dependent leak of soluble and membrane proteins through the yeast nuclear pore complex
Abstract
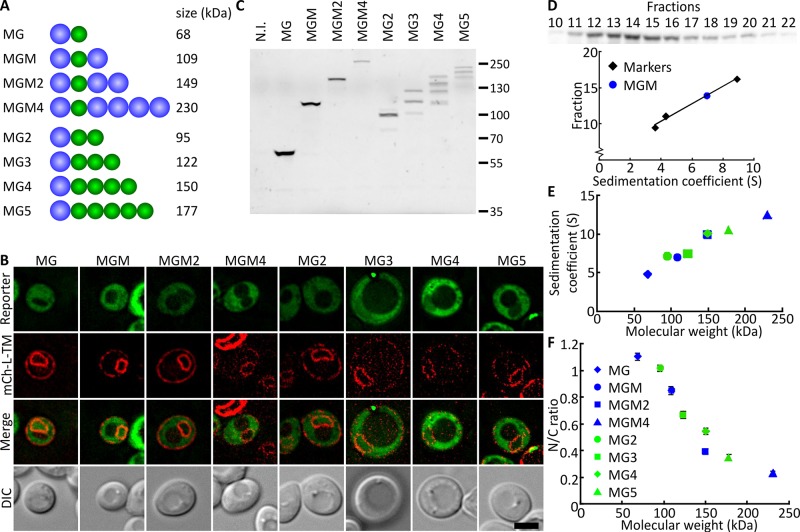
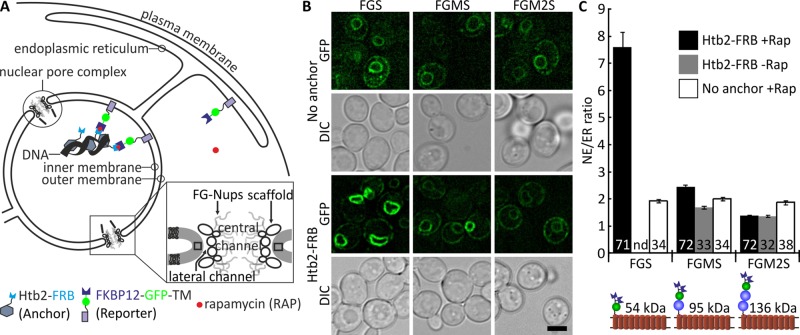
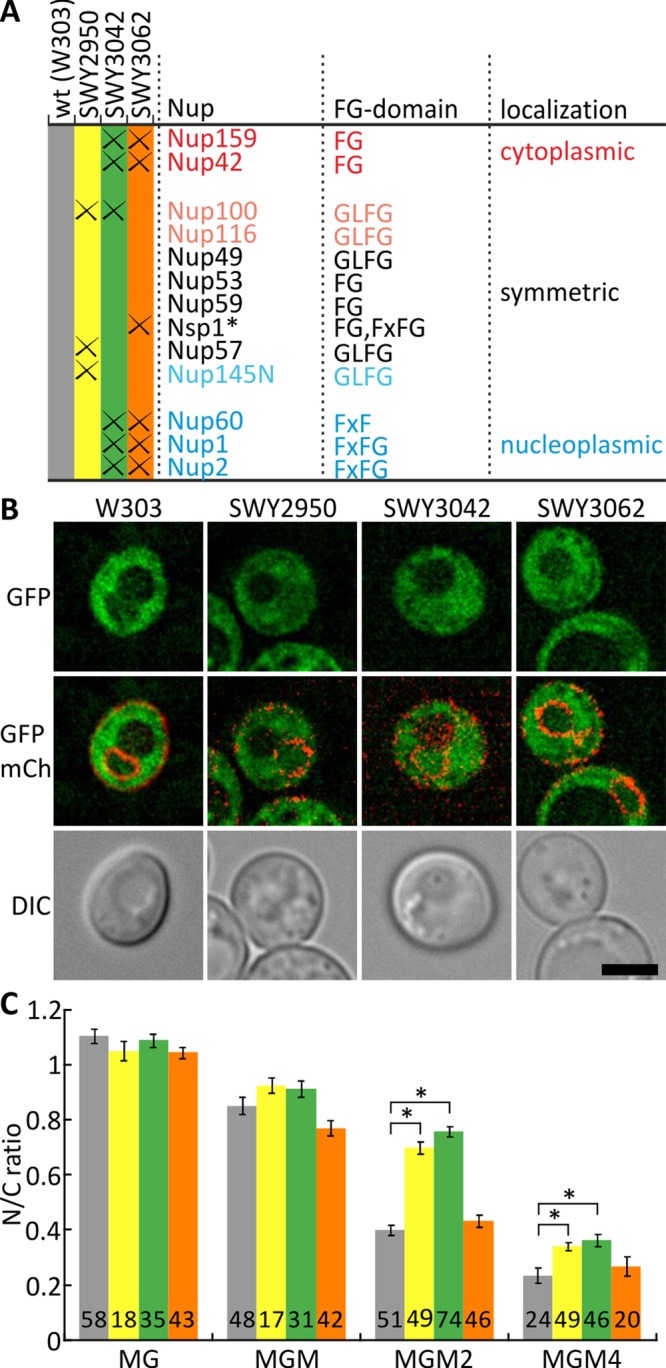
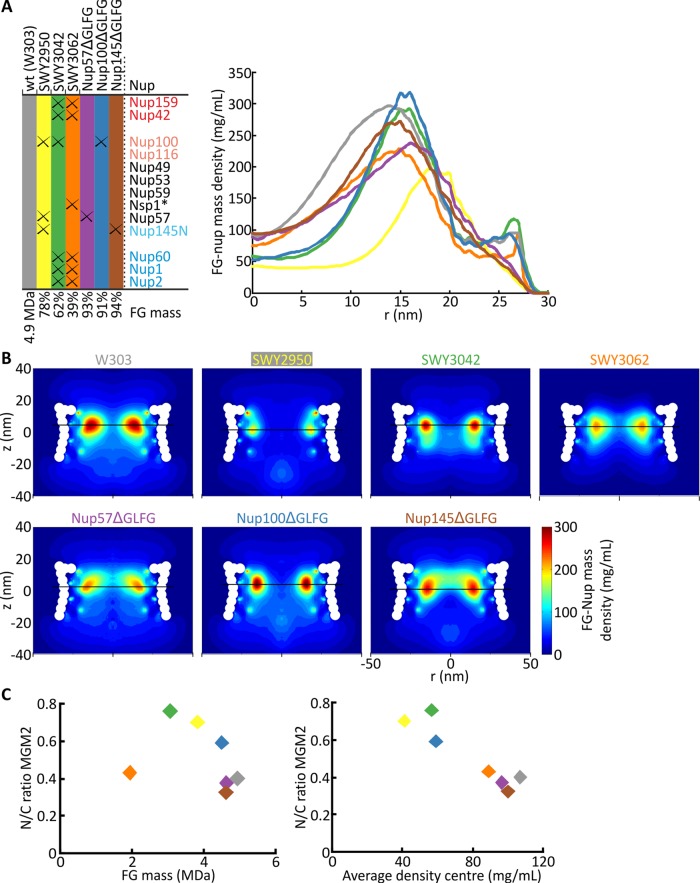
Nuclear pore complexes (NPCs) allow selective import and export while forming a barrier for untargeted proteins. Using fluorescence microscopy, we measured in vivo the permeability of the Saccharomyces cerevisiae NPC for multidomain proteins of different sizes and found that soluble proteins of 150 kDa and membrane proteins with an extralumenal domain of 90 kDa were still partly localized in the nucleus on a time scale of hours. The NPCs thus form only a weak barrier for the majority of yeast proteins, given their monomeric size. Using FGΔ-mutant strains, we showed that specific combinations of Nups, especially with Nup100, but not the total mass of FG-nups per pore, were important for forming the barrier. Models of the disordered phase of wild-type and mutant NPCs were generated using a one bead per amino acid molecular dynamics model. The permeability measurements correlated with the density predictions from coarse-grained molecular dynamics simulations in the center of the NPC. The combined in vivo and computational approach provides a framework for elucidating the structural and functional properties of the permeability barrier of nuclear pore complexes.
© 2015 Popken et al. This article is distributed by The American Society for Cell Biology under license from the author(s). Two months after publication it is available to the public under an Attribution–Noncommercial–Share Alike 3.0 Unported Creative Commons License (http://creativecommons.org/licenses/by-nc-sa/3.0).
Figures
References
-
- Alber F, Dokudovskaya S, Veenhoff LM, Zhang W, Kipper J, Devos D, Suprapto A, Karni-Schmidt O, Williams R, Chait BT, et al. The molecular architecture of the nuclear pore complex. Nature. 2007;450:695–701. - PubMed
-
- Beetz C, Brodhun M, Moutzouris K, Kiehntopf M, Berndt A, Lehnert D, Deufel T, Bastmeyer M, Schickel J. Identification of nuclear localisation sequences in spastin (SPG4) using a novel Tetra-GFP reporter system. Biochem Biophys Res Commun. 2004;318:1079–1084. - PubMed
-
- Chatterjee S, Javier M, Stochaj U. In vivo analysis of nuclear protein traffic in mammalian cells. Exp Cell Res. 1997;236:346–350. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
