

Dysfunction in the βII spectrin-dependent cytoskeleton underlies human arrhythmia
- PMID: 25632041
- PMCID: PMC4342332
- DOI: 10.1161/CIRCULATIONAHA.114.013708
Dysfunction in the βII spectrin-dependent cytoskeleton underlies human arrhythmia
Abstract
Background: The cardiac cytoskeleton plays key roles in maintaining myocyte structural integrity in health and disease. In fact, human mutations in cardiac cytoskeletal elements are tightly linked to cardiac pathologies, including myopathies, aortopathies, and dystrophies. Conversely, the link between cytoskeletal protein dysfunction and cardiac electric activity is not well understood and often overlooked in the cardiac arrhythmia field.
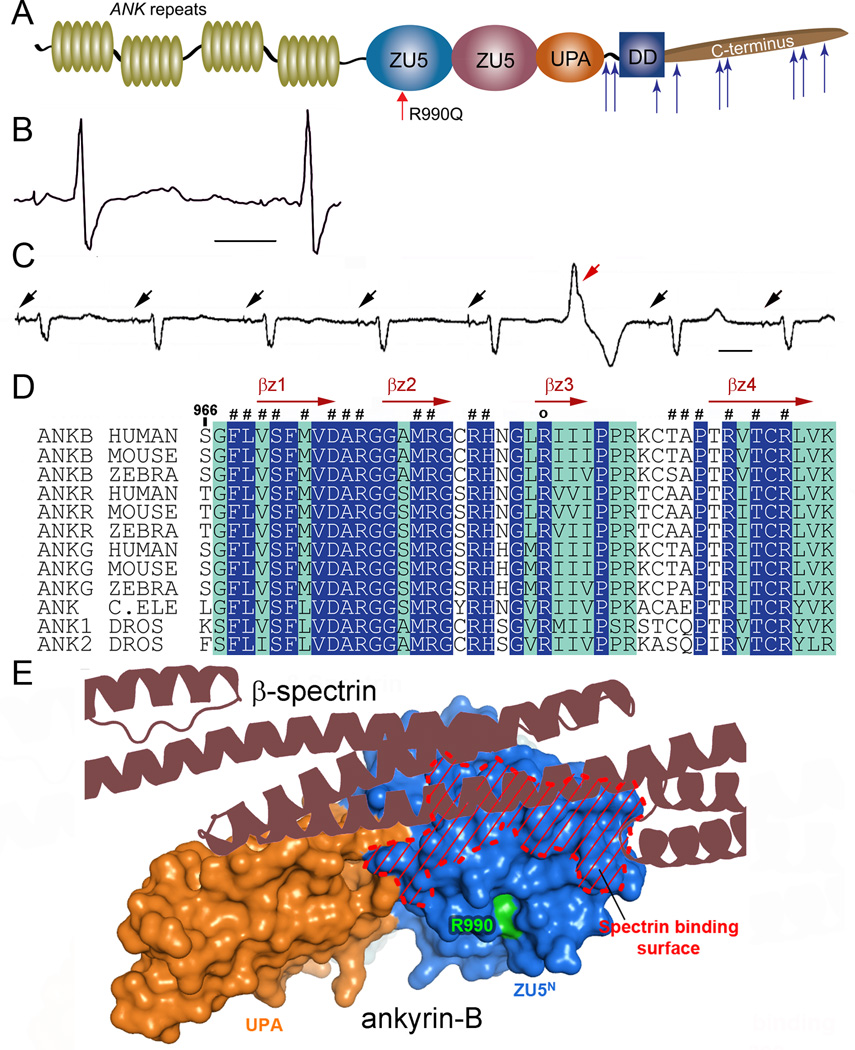
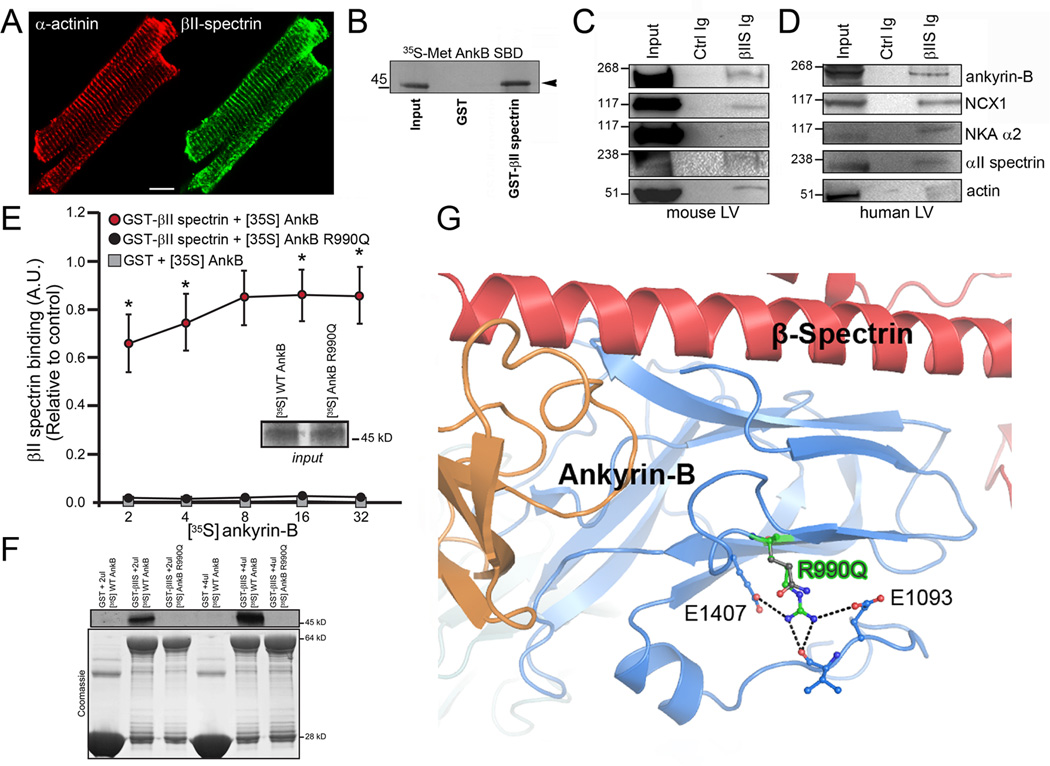
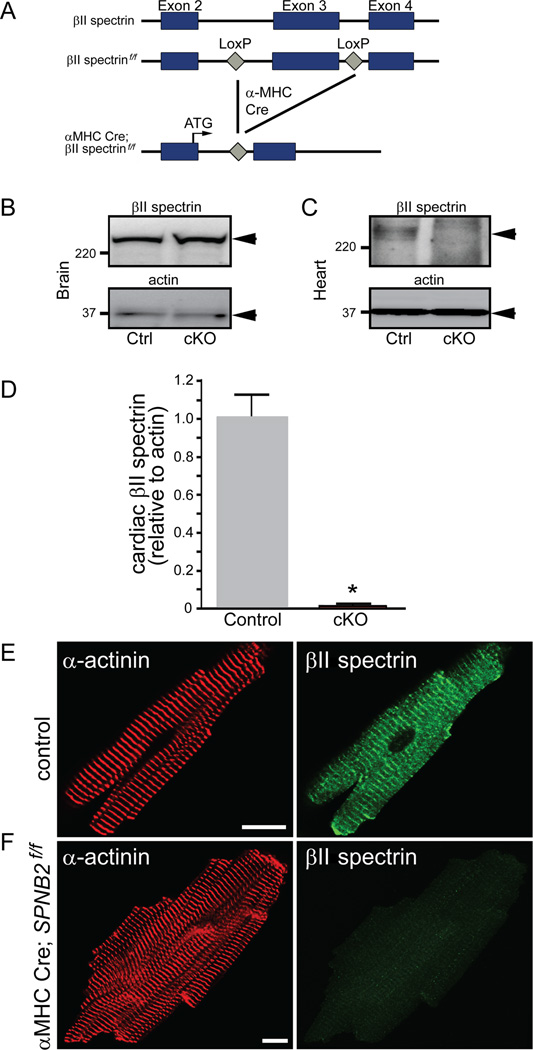
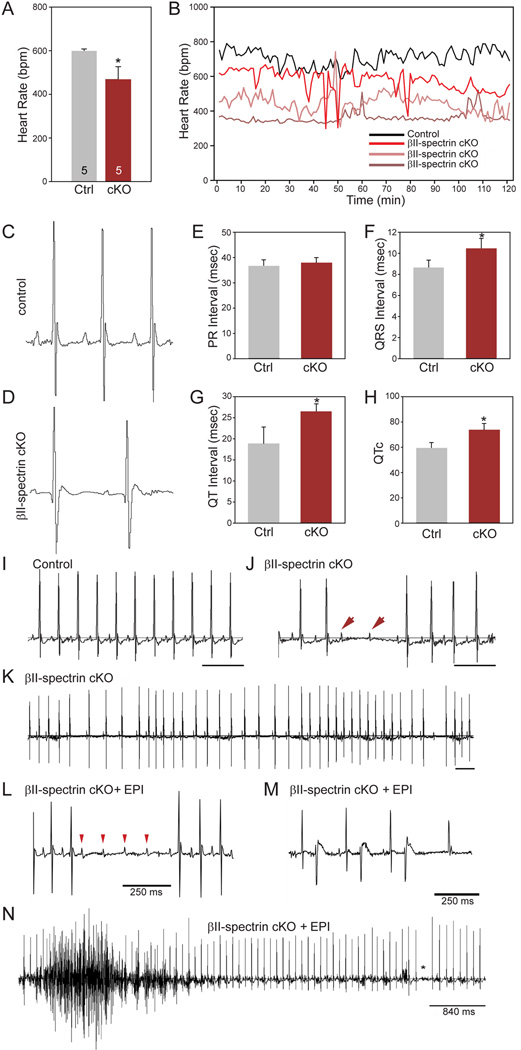
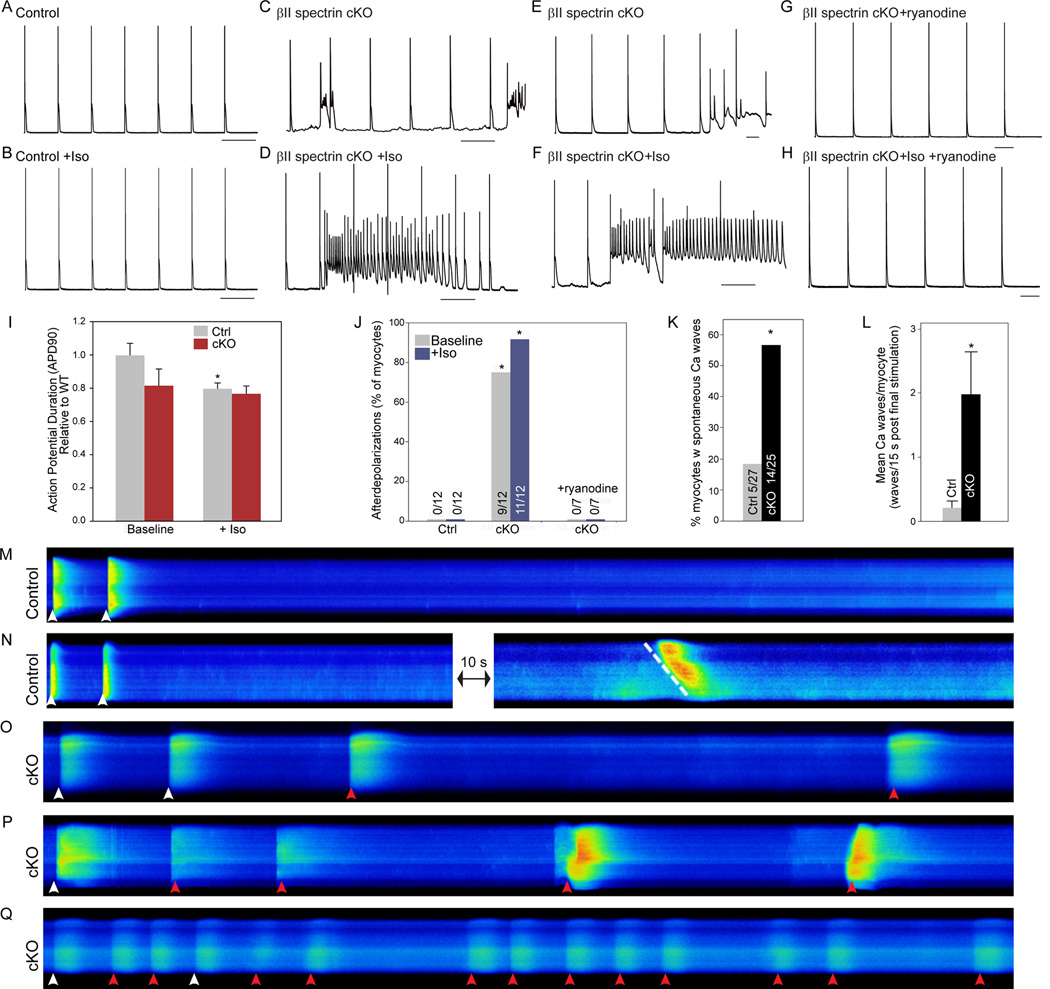
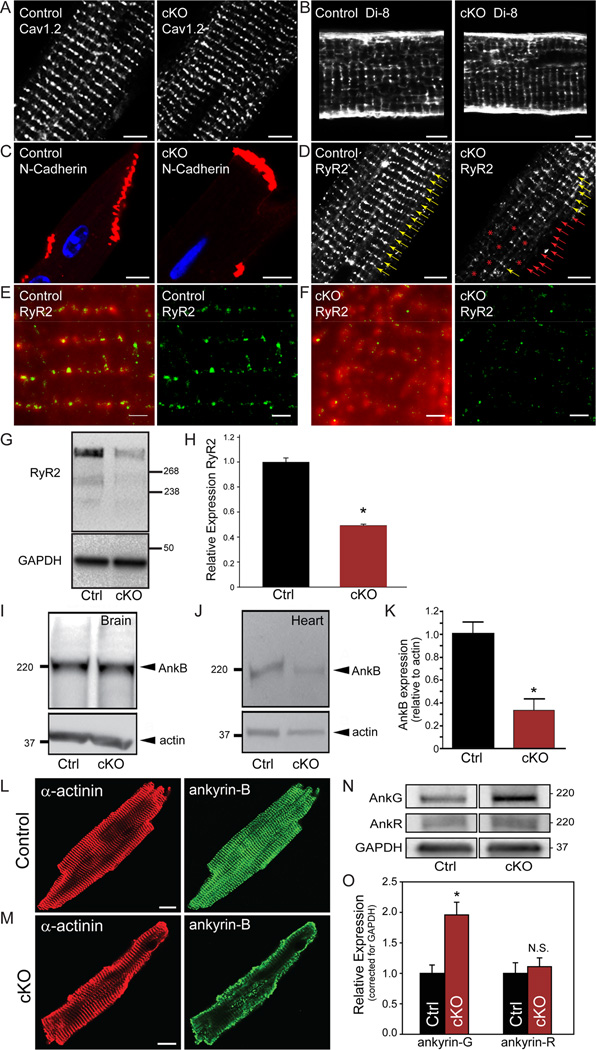
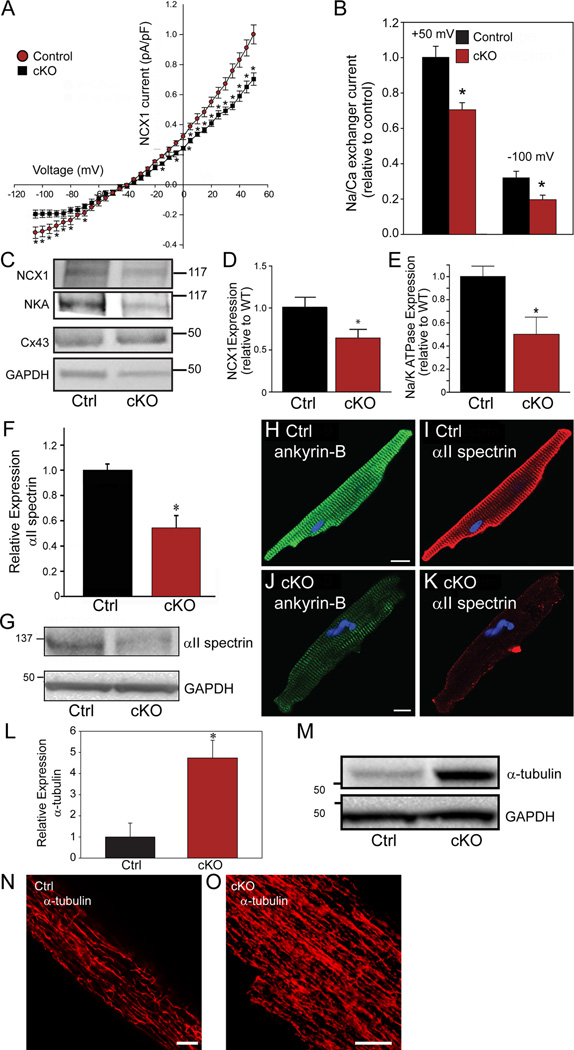
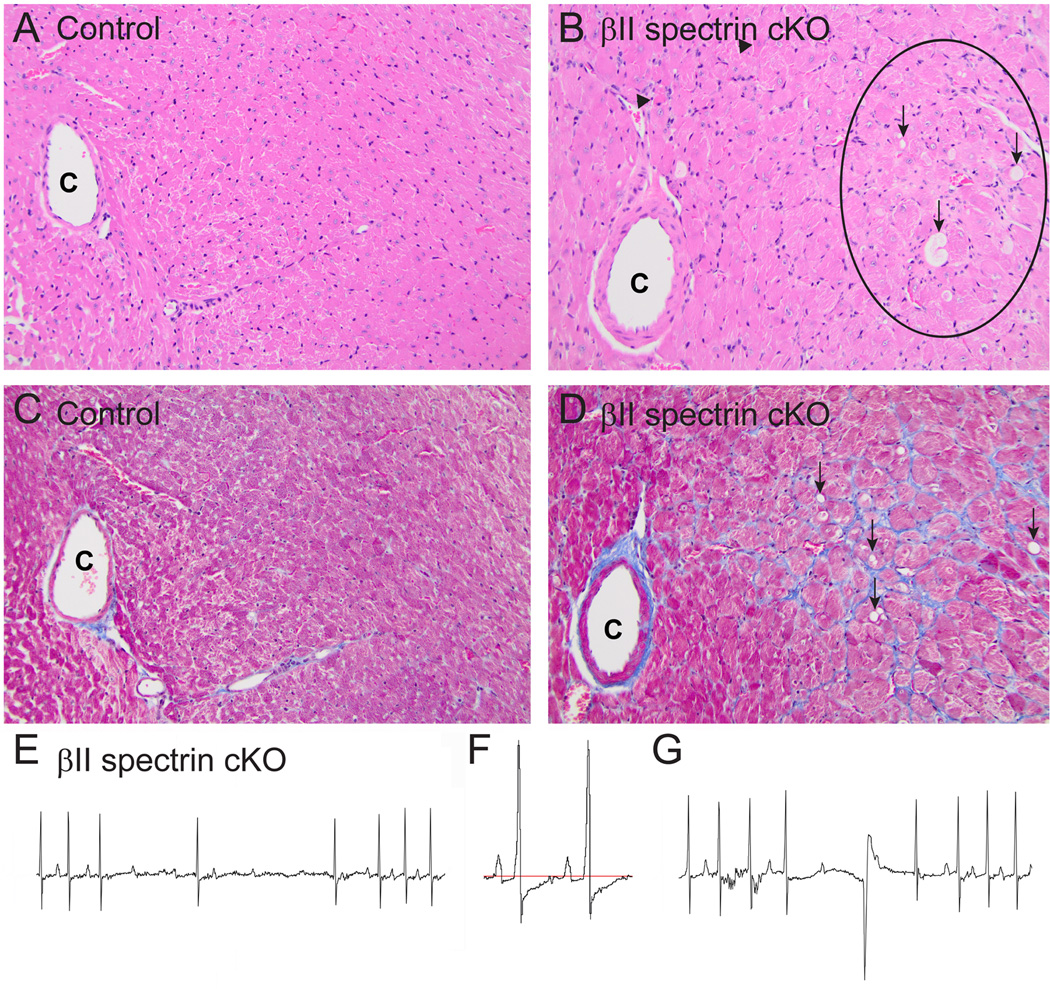
Methods and results: Here, we uncover a new mechanism for the regulation of cardiac membrane excitability. We report that βII spectrin, an actin-associated molecule, is essential for the posttranslational targeting and localization of critical membrane proteins in heart. βII spectrin recruits ankyrin-B to the cardiac dyad, and a novel human mutation in the ankyrin-B gene disrupts the ankyrin-B/βII spectrin interaction, leading to severe human arrhythmia phenotypes. Mice lacking cardiac βII spectrin display lethal arrhythmias, aberrant electric and calcium handling phenotypes, and abnormal expression/localization of cardiac membrane proteins. Mechanistically, βII spectrin regulates the localization of cytoskeletal and plasma membrane/sarcoplasmic reticulum protein complexes, including the Na/Ca exchanger, ryanodine receptor 2, ankyrin-B, actin, and αII spectrin. Finally, we observe accelerated heart failure phenotypes in βII spectrin-deficient mice.
Conclusions: Our findings identify βII spectrin as critical for normal myocyte electric activity, link this molecule to human disease, and provide new insight into the mechanisms underlying cardiac myocyte biology.
Keywords: arrhythmias, cardiac; catecholaminergic polymorphic ventricular arrhythmia; cytoskeleton; ion channels; protein transport; ventricular tachycardia.
© 2015 American Heart Association, Inc.
Figures
Comment in
-
Cytoskeleton regulation of ion channels.Circulation. 2015 Feb 24;131(8):689-91. doi: 10.1161/CIRCULATIONAHA.115.015216. Epub 2015 Jan 28. Circulation. 2015. PMID: 25632042 Free PMC article. No abstract available.
References
-
- van Tintelen JP, Entius MM, Bhuiyan ZA, Jongbloed R, Wiesfeld AC, Wilde AA, van der Smagt J, Boven LG, Mannens MM, van Langen IM, Hofstra RM, Otterspoor LC, Doevendans PA, Rodriguez LM, van Gelder IC, Hauer RN. Plakophilin-2 mutations are the major determinant of familial arrhythmogenic right ventricular dysplasia/cardiomyopathy. Circulation. 2006;113:1650–1658. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
