

Cellular origin and developmental program of coronary angiogenesis
- PMID: 25634974
- PMCID: PMC6914229
- DOI: 10.1161/CIRCRESAHA.116.305097
Cellular origin and developmental program of coronary angiogenesis
Abstract
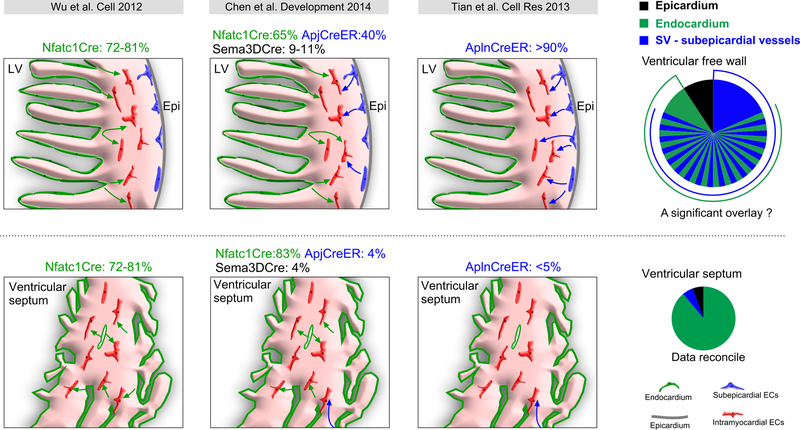
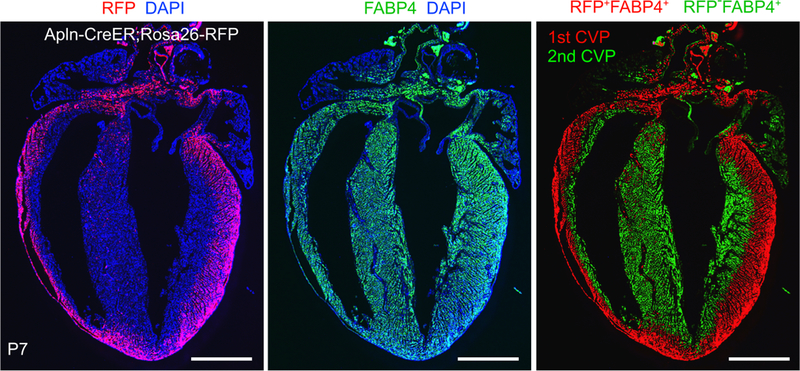
Coronary artery disease causes acute myocardial infarction and heart failure. Identifying coronary vascular progenitors and their developmental program could inspire novel regenerative treatments for cardiac diseases. The developmental origins of the coronary vessels have been shrouded in mystery and debated for several decades. Recent identification of progenitors for coronary vessels within the endocardium, epicardium, and sinus venosus provides new insights into this question. In addition, significant progress has been achieved in elucidating the cellular and molecular programs that orchestrate coronary artery development. Establishing adequate vascular supply will be an essential component of cardiac regenerative strategies, and these findings raise exciting new strategies for therapeutic cardiac revascularization.
Keywords: coronary vessels; endocardium; sinus venosus.
© 2015 American Heart Association, Inc.
Figures
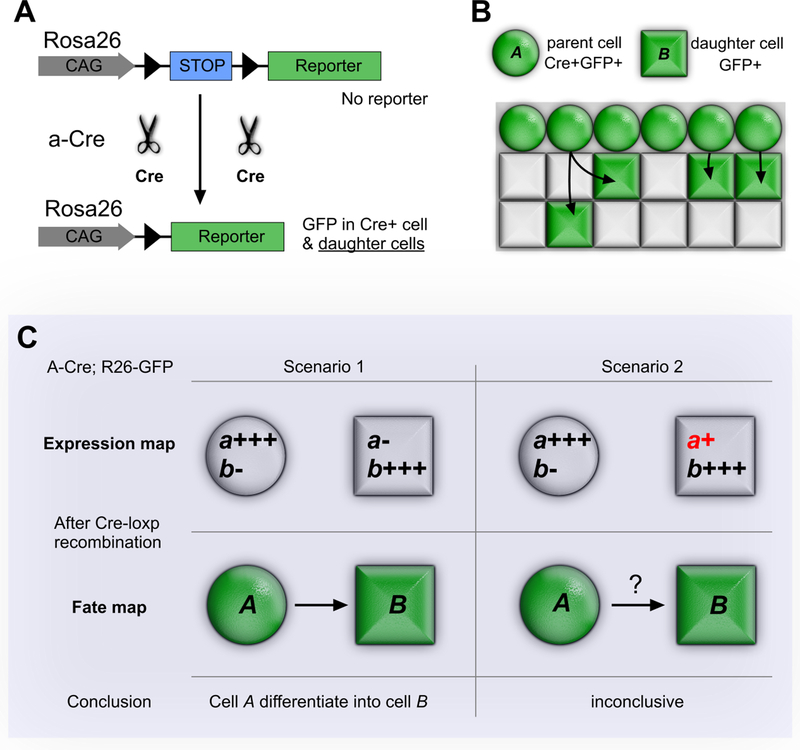
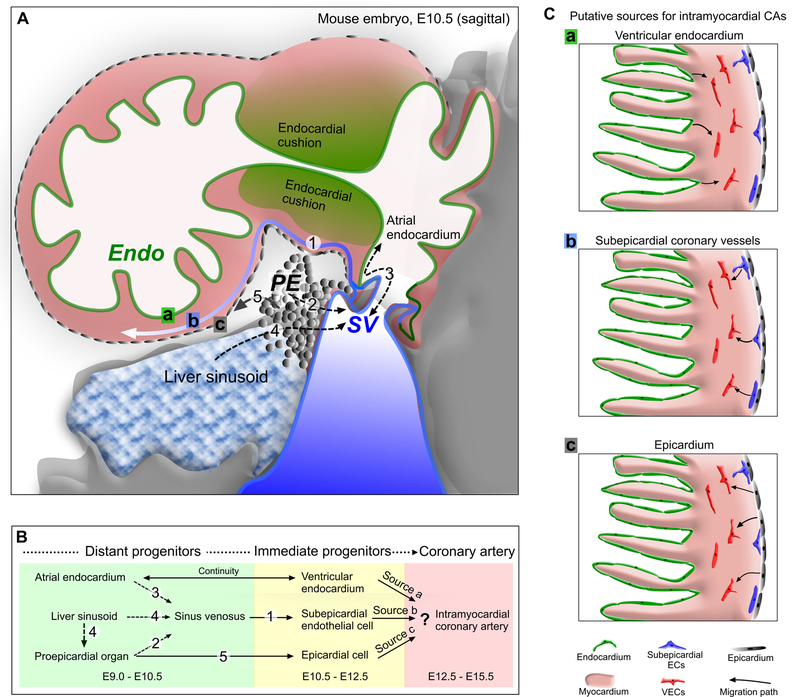
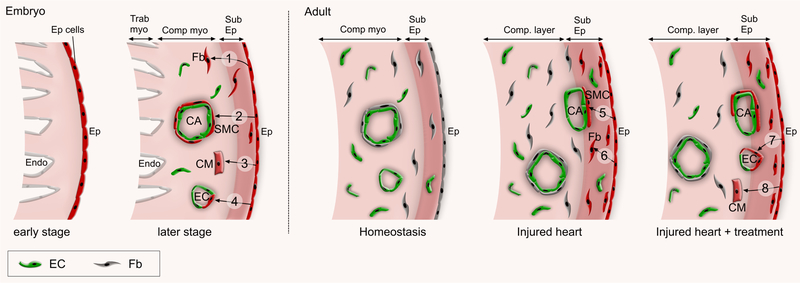
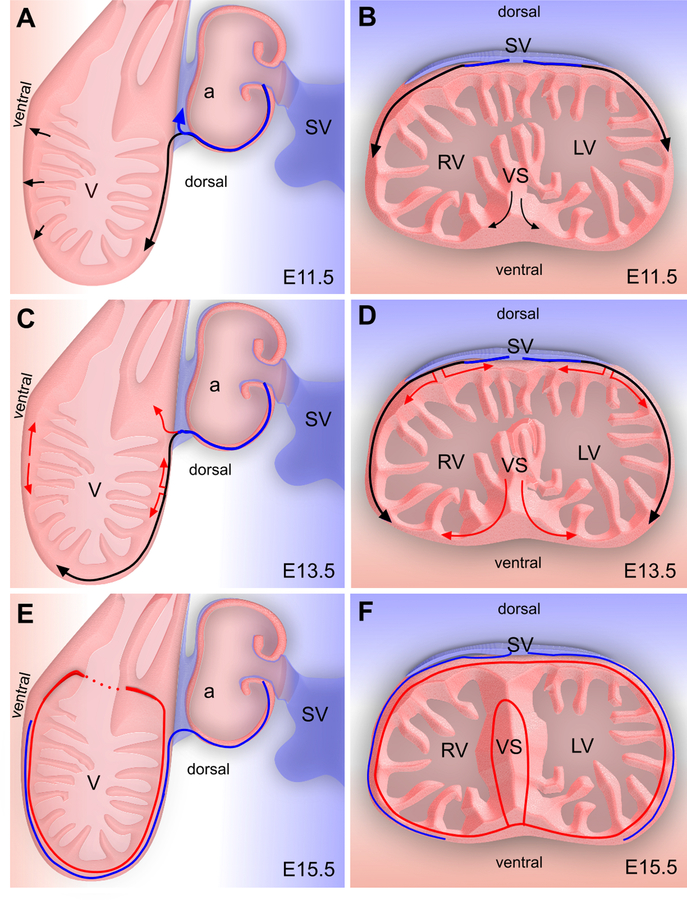
References
-
- Hansson EM, Lindsay ME, Chien KR. Regeneration next: toward heart stem cell therapeutics. Cell Stem Cell 2009;5:364–377. - PubMed
-
- Martin-Puig S, Wang Z, Chien KR. Lives of a heart cell: tracing the origins of cardiac progenitors. Cell Stem Cell 2008;2:320–331. - PubMed
-
- Zhou B, Lin Z, Pu W. Mammalian Myocardial Regeneration. Muscle 2012;First Edition, Chapter 39:555–569.
-
- Passier R, van Laake LW, Mummery CL. Stem-cell-based therapy and lessons from the heart. Nature 2008;453:322–329. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
