

Perturbation of maize phenylpropanoid metabolism by an AvrE family type III effector from Pantoea stewartii
- PMID: 25635112
- PMCID: PMC4348765
- DOI: 10.1104/pp.114.253120
Perturbation of maize phenylpropanoid metabolism by an AvrE family type III effector from Pantoea stewartii
Abstract
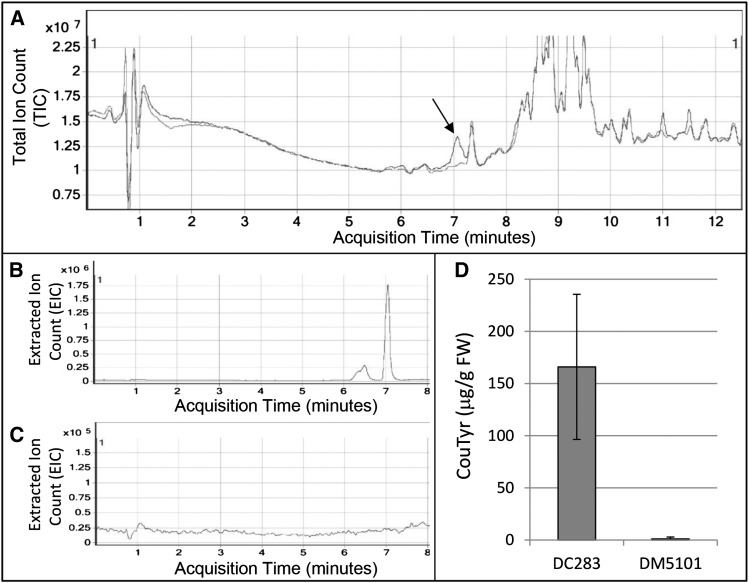
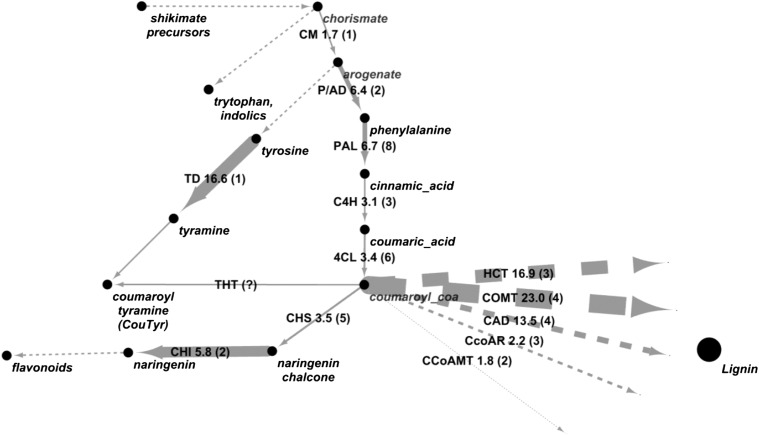
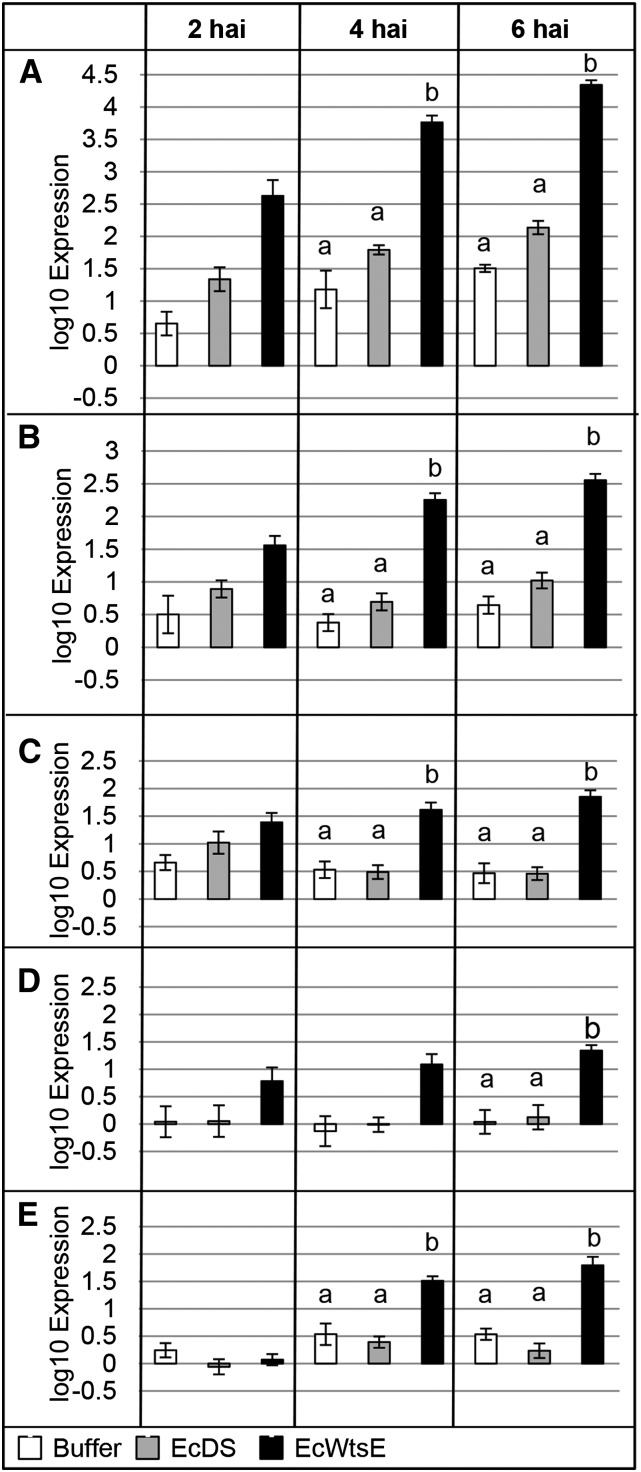
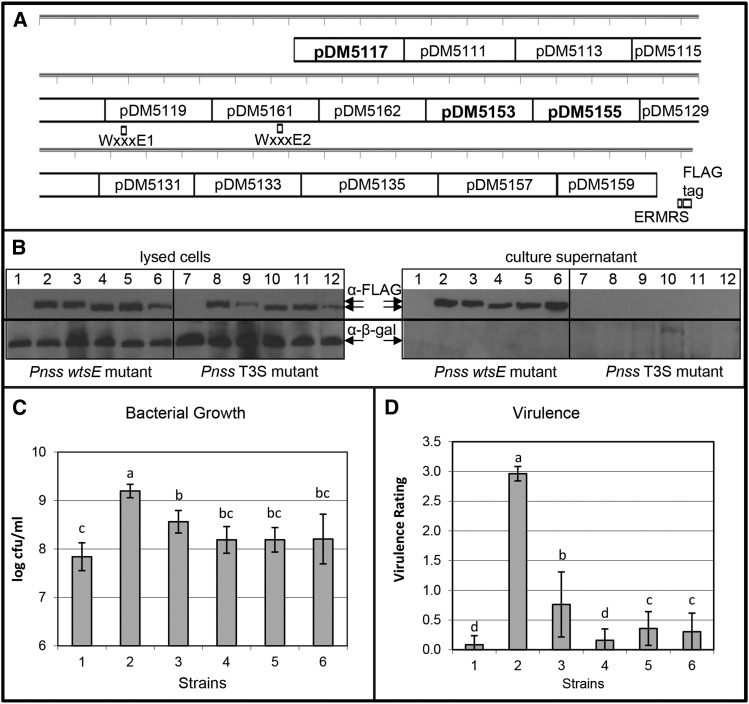
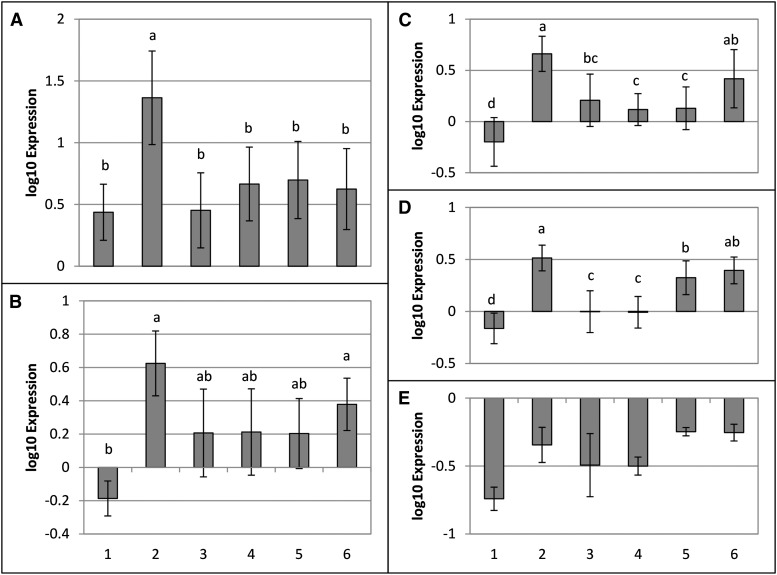
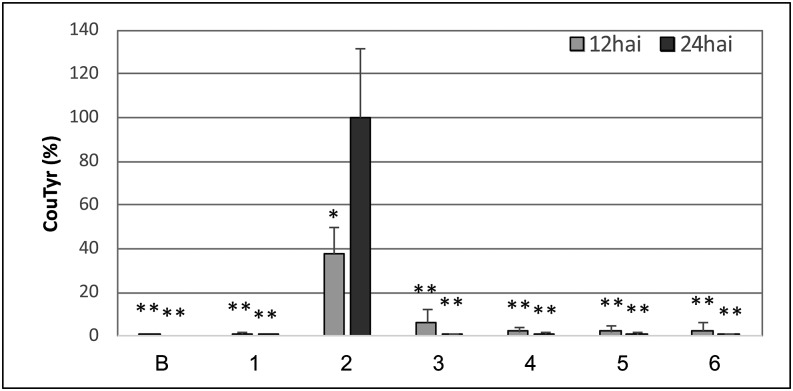
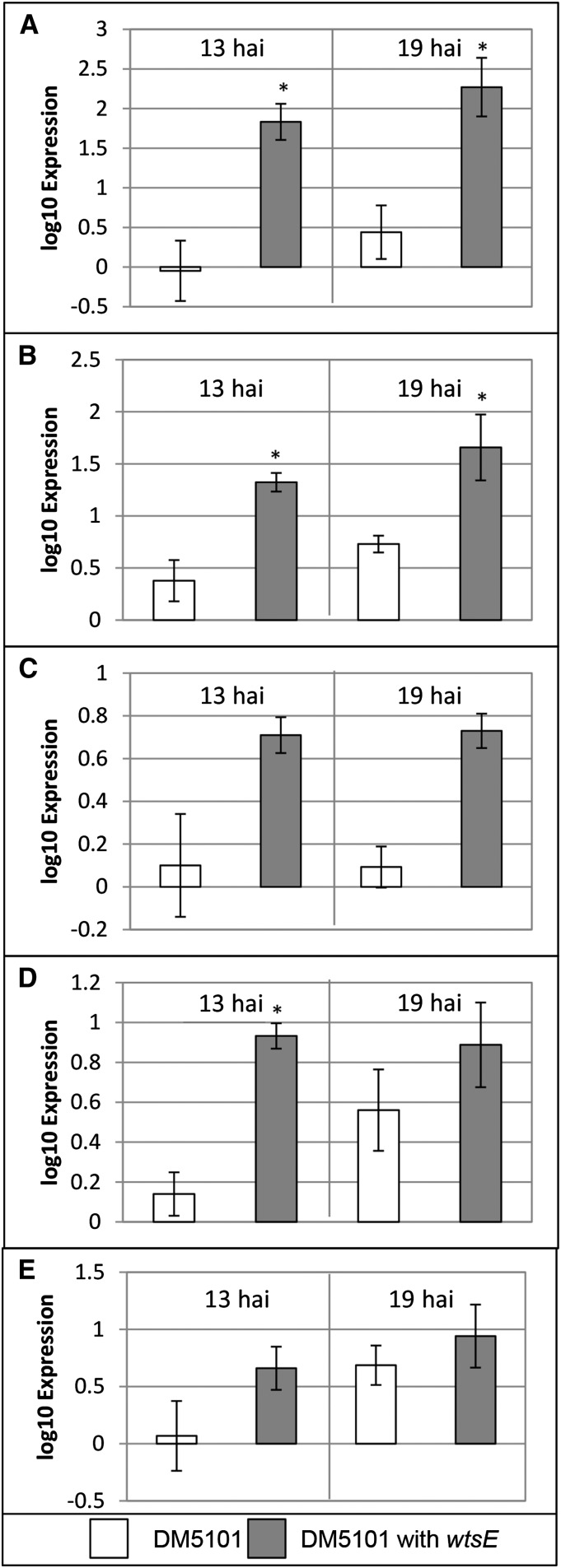
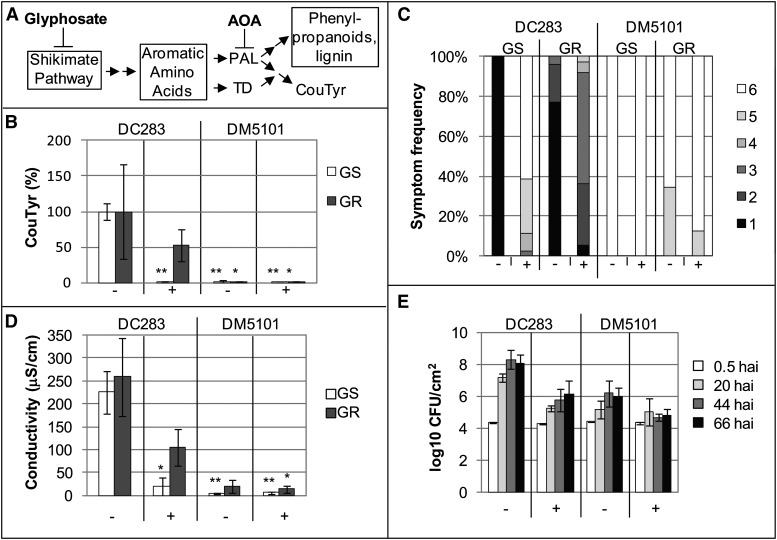
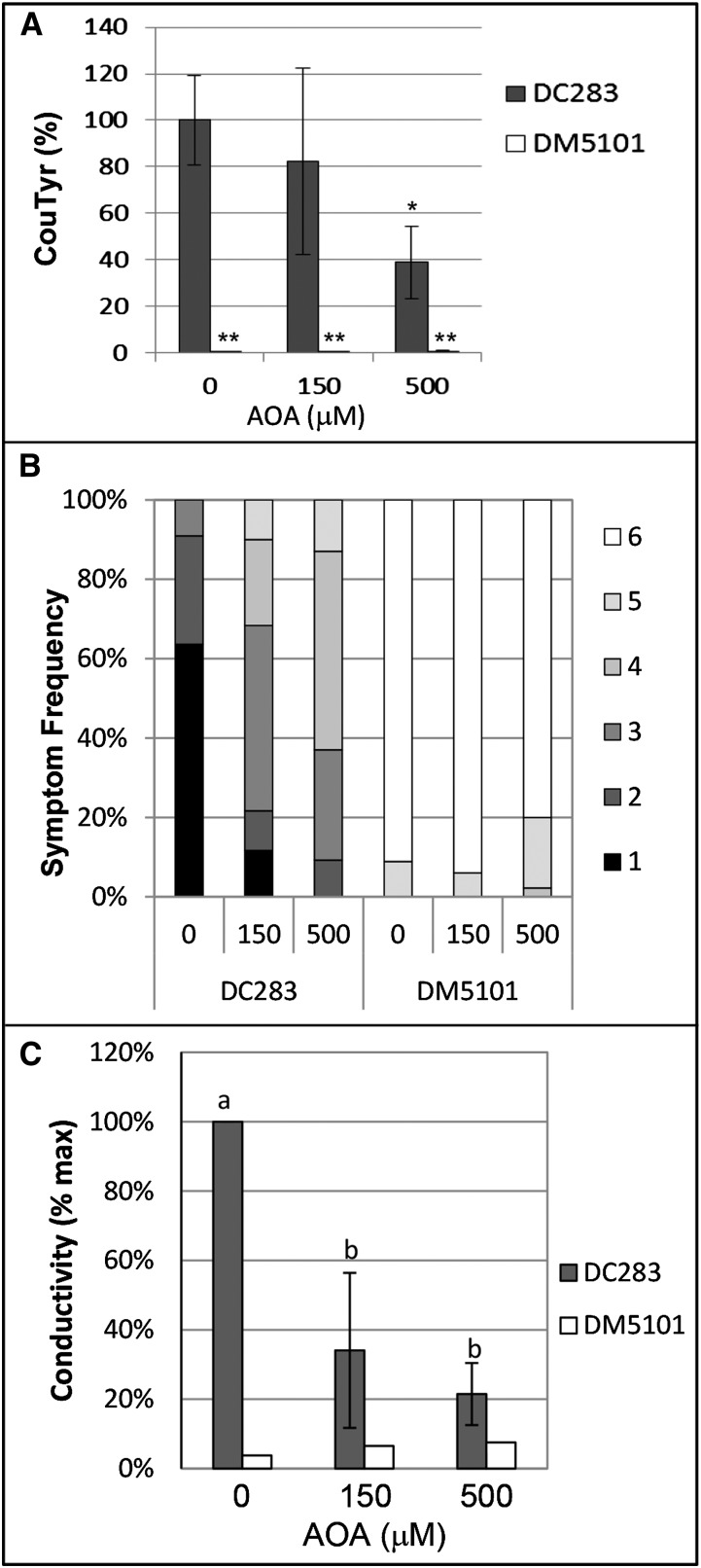
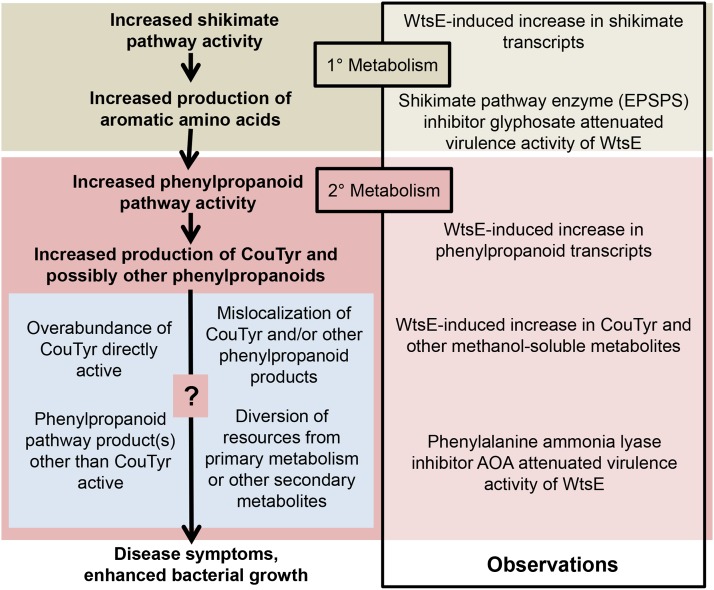
AvrE family type III effector proteins share the ability to suppress host defenses, induce disease-associated cell death, and promote bacterial growth. However, despite widespread contributions to numerous bacterial diseases in agriculturally important plants, the mode of action of these effectors remains largely unknown. WtsE is an AvrE family member required for the ability of Pantoea stewartii ssp. stewartii (Pnss) to proliferate efficiently and cause wilt and leaf blight symptoms in maize (Zea mays) plants. Notably, when WtsE is delivered by a heterologous system into the leaf cells of susceptible maize seedlings, it alone produces water-soaked disease symptoms reminiscent of those produced by Pnss. Thus, WtsE is a pathogenicity and virulence factor in maize, and an Escherichia coli heterologous delivery system can be used to study the activity of WtsE in isolation from other factors produced by Pnss. Transcriptional profiling of maize revealed the effects of WtsE, including induction of genes involved in secondary metabolism and suppression of genes involved in photosynthesis. Targeted metabolite quantification revealed that WtsE perturbs maize metabolism, including the induction of coumaroyl tyramine. The ability of mutant WtsE derivatives to elicit transcriptional and metabolic changes in susceptible maize seedlings correlated with their ability to promote disease. Furthermore, chemical inhibitors that block metabolic flux into the phenylpropanoid pathways targeted by WtsE also disrupted the pathogenicity and virulence activity of WtsE. While numerous metabolites produced downstream of the shikimate pathway are known to promote plant defense, our results indicate that misregulated induction of phenylpropanoid metabolism also can be used to promote pathogen virulence.
© 2015 American Society of Plant Biologists. All Rights Reserved.
Figures
References
-
- Ahmad M, Majerczak DR, Pike S, Hoyos ME, Novacky A, Coplin DL (2001) Biological activity of harpin produced by Pantoea stewartii subsp. stewartii. Mol Plant Microbe Interact 14: 1223–1234 - PubMed
-
- Ahuja I, Kissen R, Bones AM (2012) Phytoalexins in defense against pathogens. Trends Plant Sci 17: 73–90 - PubMed
-
- Alto NM, Shao F, Lazar CS, Brost RL, Chua G, Mattoo S, McMahon SA, Ghosh P, Hughes TR, Boone C, et al. (2006) Identification of a bacterial type III effector family with G protein mimicry functions. Cell 124: 133–145 - PubMed
-
- Ausubel FM, Brent R, Kingston RE, More DD, Seidman JG, Smith JA, Struhl K (1997) Short Protocols in Molecular Biology: A Compendium of Methods from Current Protocols in Molecular Biology, Ed 3.John Wiley & Sons, New York
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
