

Decoding neural circuits that control compulsive sucrose seeking
- PMID: 25635460
- PMCID: PMC4312417
- DOI: 10.1016/j.cell.2015.01.003
Decoding neural circuits that control compulsive sucrose seeking
Abstract
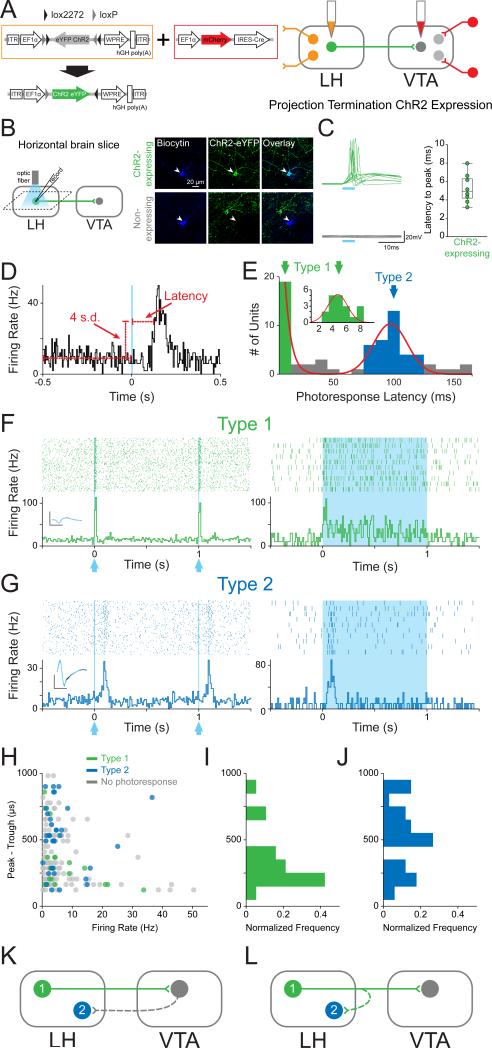
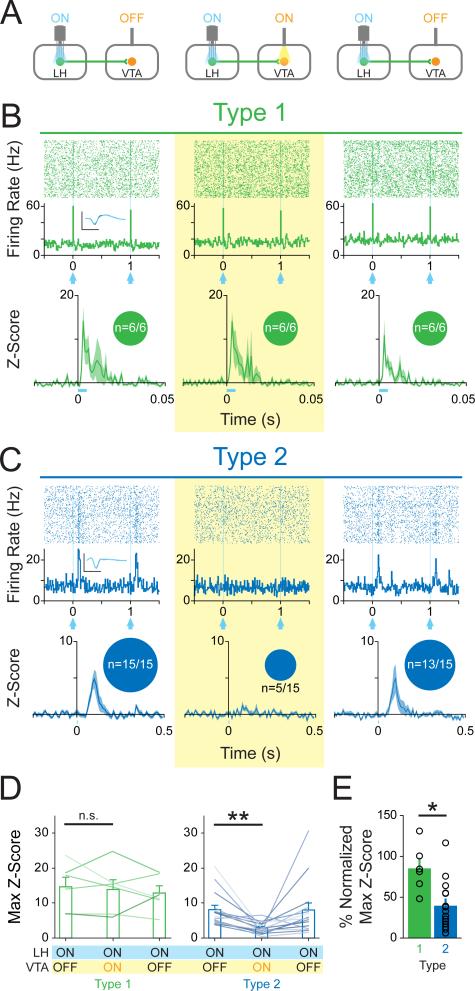
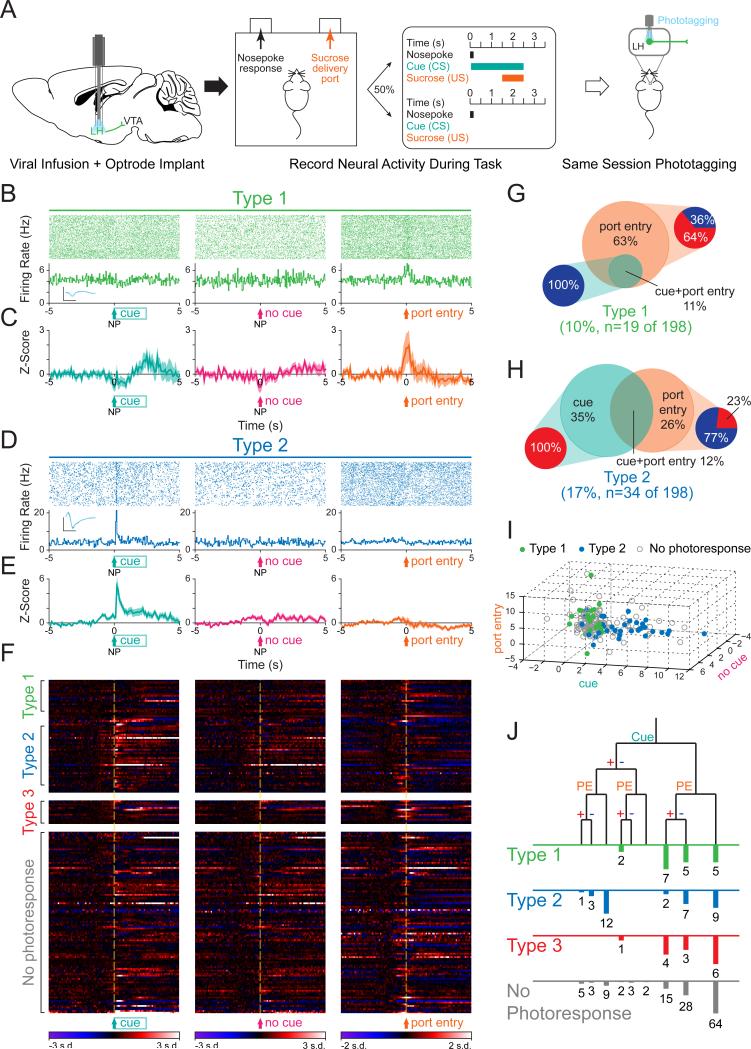
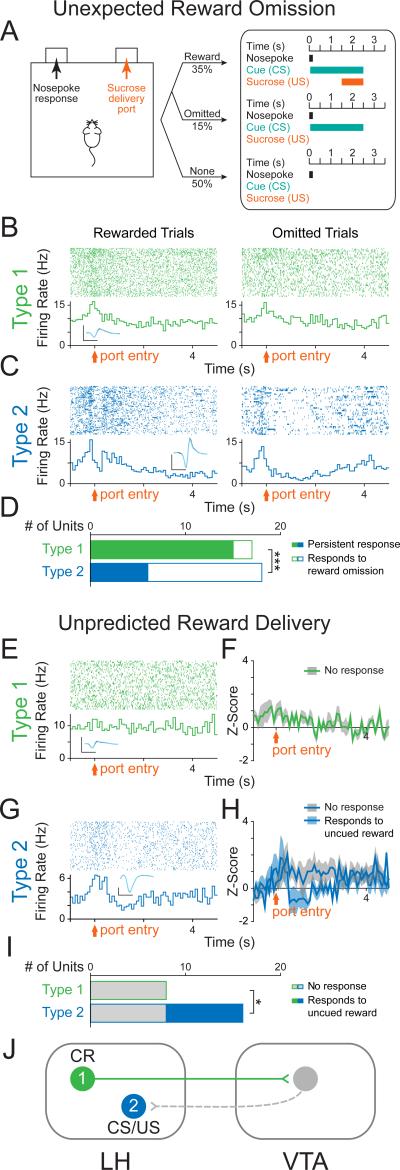
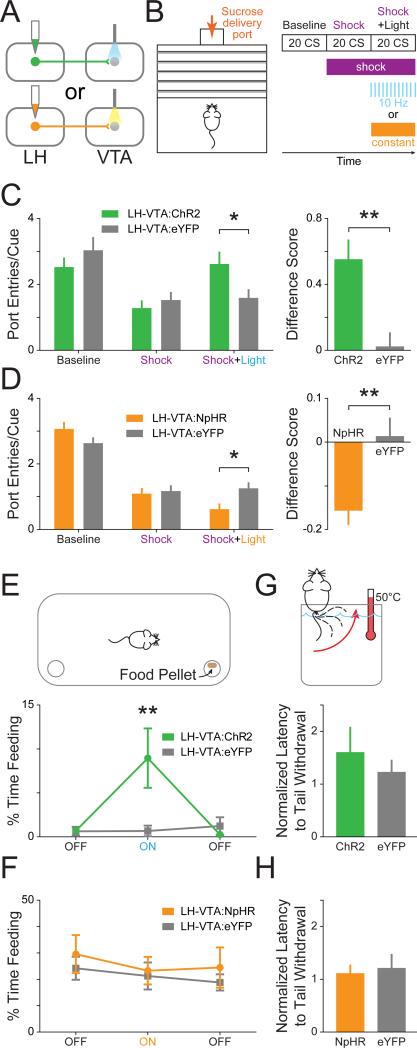
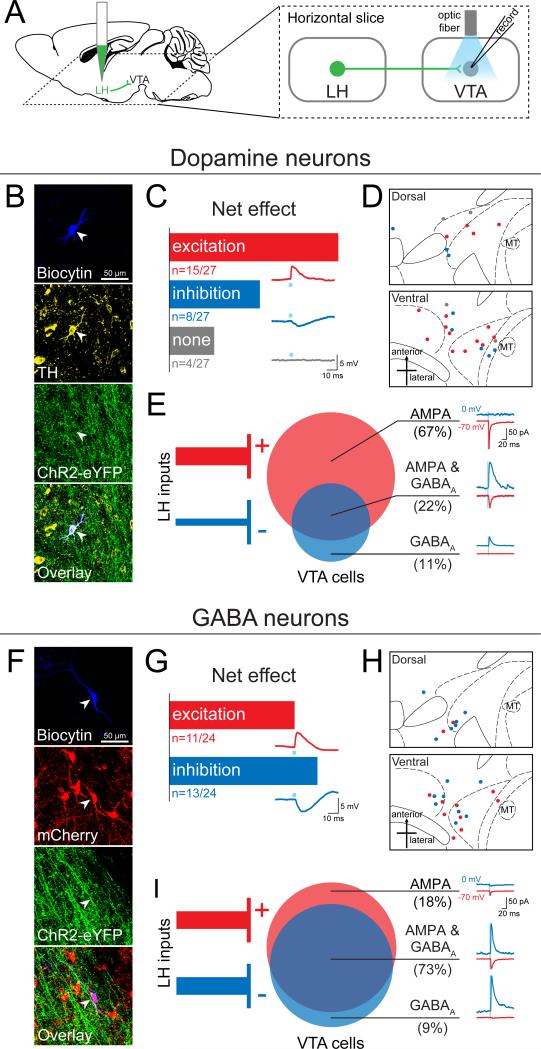
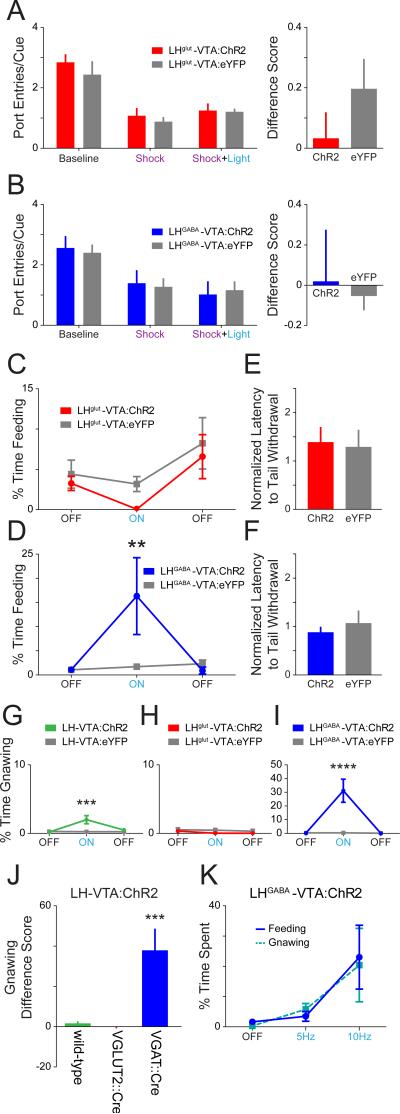
The lateral hypothalamic (LH) projection to the ventral tegmental area (VTA) has been linked to reward processing, but the computations within the LH-VTA loop that give rise to specific aspects of behavior have been difficult to isolate. We show that LH-VTA neurons encode the learned action of seeking a reward, independent of reward availability. In contrast, LH neurons downstream of VTA encode reward-predictive cues and unexpected reward omission. We show that inhibiting the LH-VTA pathway reduces "compulsive" sucrose seeking but not food consumption in hungry mice. We reveal that the LH sends excitatory and inhibitory input onto VTA dopamine (DA) and GABA neurons, and that the GABAergic projection drives feeding-related behavior. Our study overlays information about the type, function, and connectivity of LH neurons and identifies a neural circuit that selectively controls compulsive sugar consumption, without preventing feeding necessary for survival, providing a potential target for therapeutic interventions for compulsive-overeating disorder.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Obesity. Unravelling the neural networks that drive different eating behaviours.Nat Rev Endocrinol. 2015 Apr;11(4):191. doi: 10.1038/nrendo.2015.16. Epub 2015 Feb 17. Nat Rev Endocrinol. 2015. PMID: 25687992 No abstract available.
References
-
- Avena NM. Examining the addictive-like properties of binge eating using an animal model of sugar dependence. Experimental and Clinical Psychopharmacology; Experimental and Clinical Psychopharmacology. 2007;15:481. - PubMed
-
- Barone FC, Wayner MJ, Scharoun SL, Guevara-Aguilar R, Aguilar-Baturoni HU. Afferent connections to the lateral hypothalamus: a horseradish peroxidase study in the rat. Brain Res. Bull. 1981;7:75–88. - PubMed
-
- Ben-Bassat J, Peretz E, Sulman FG. Analgesimetry and ranking of analgesic drugs by the receptacle method. Arch Int Pharmacodyn Ther. 1959;122:434–447. - PubMed
-
- Beckstead RM, Domesick VB, Nauta WJ. Efferent connections of the substantia nigra and ventral tegmental area in the rat. Brain Res. 1979;175:191–217. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
