

Mistakes were made: neural mechanisms for the adaptive control of action initiation by the medial prefrontal cortex
- PMID: 25636373
- PMCID: PMC5292776
- DOI: 10.1016/j.jphysparis.2014.12.001
Mistakes were made: neural mechanisms for the adaptive control of action initiation by the medial prefrontal cortex
Abstract
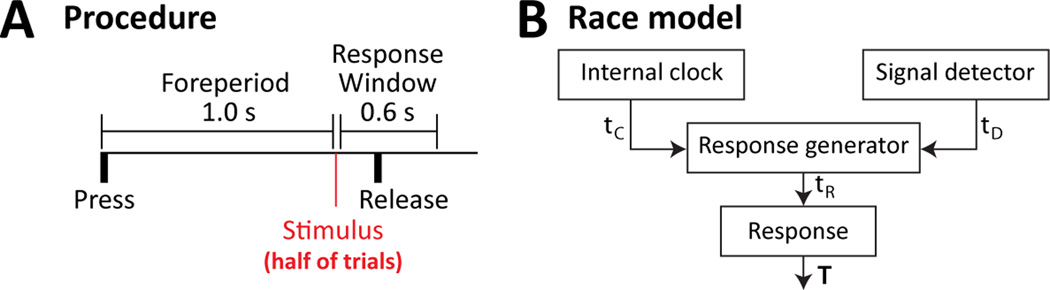
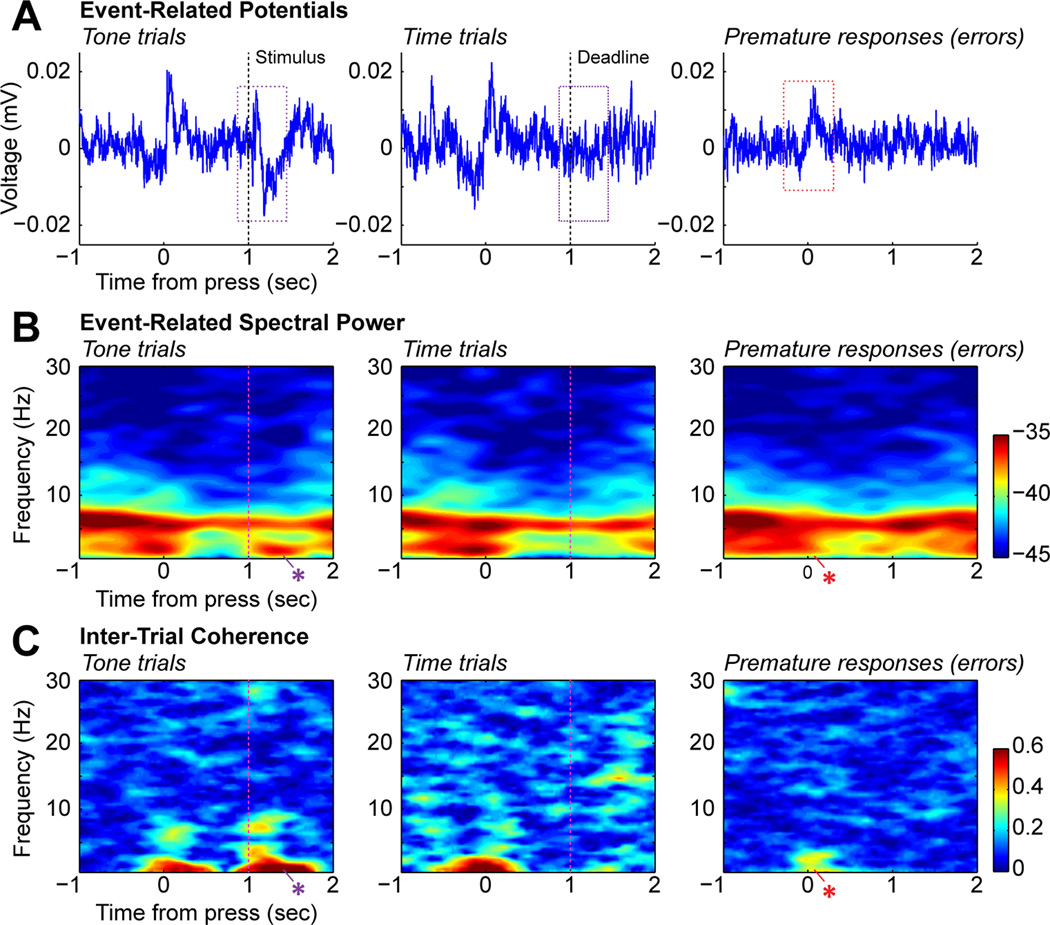
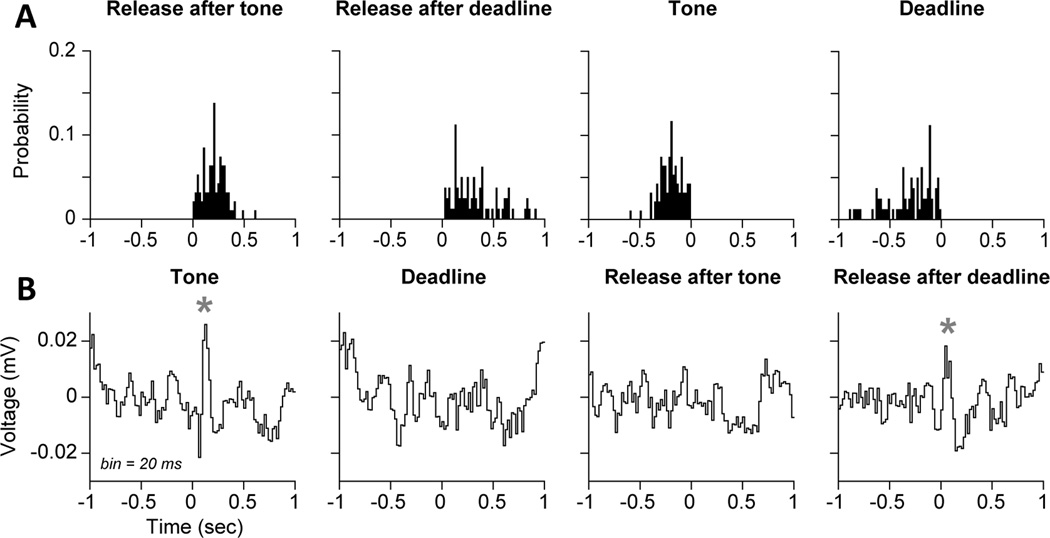
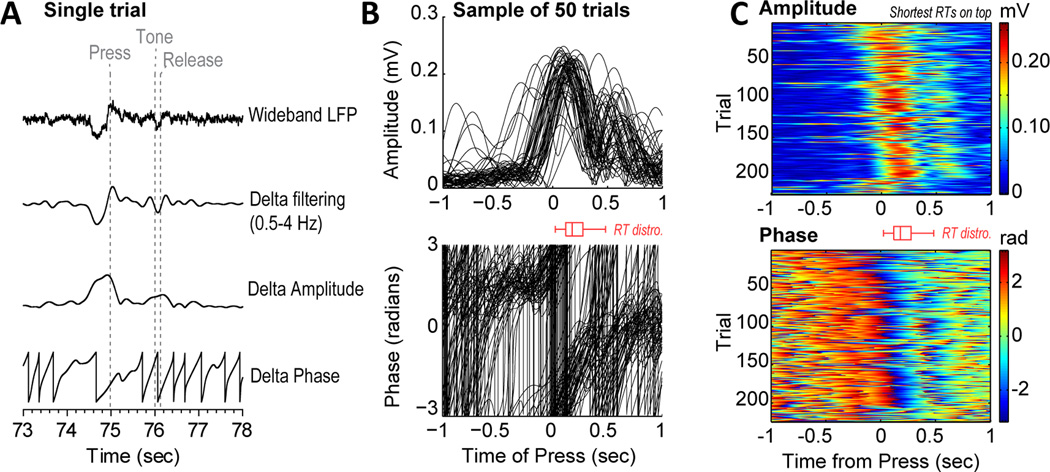
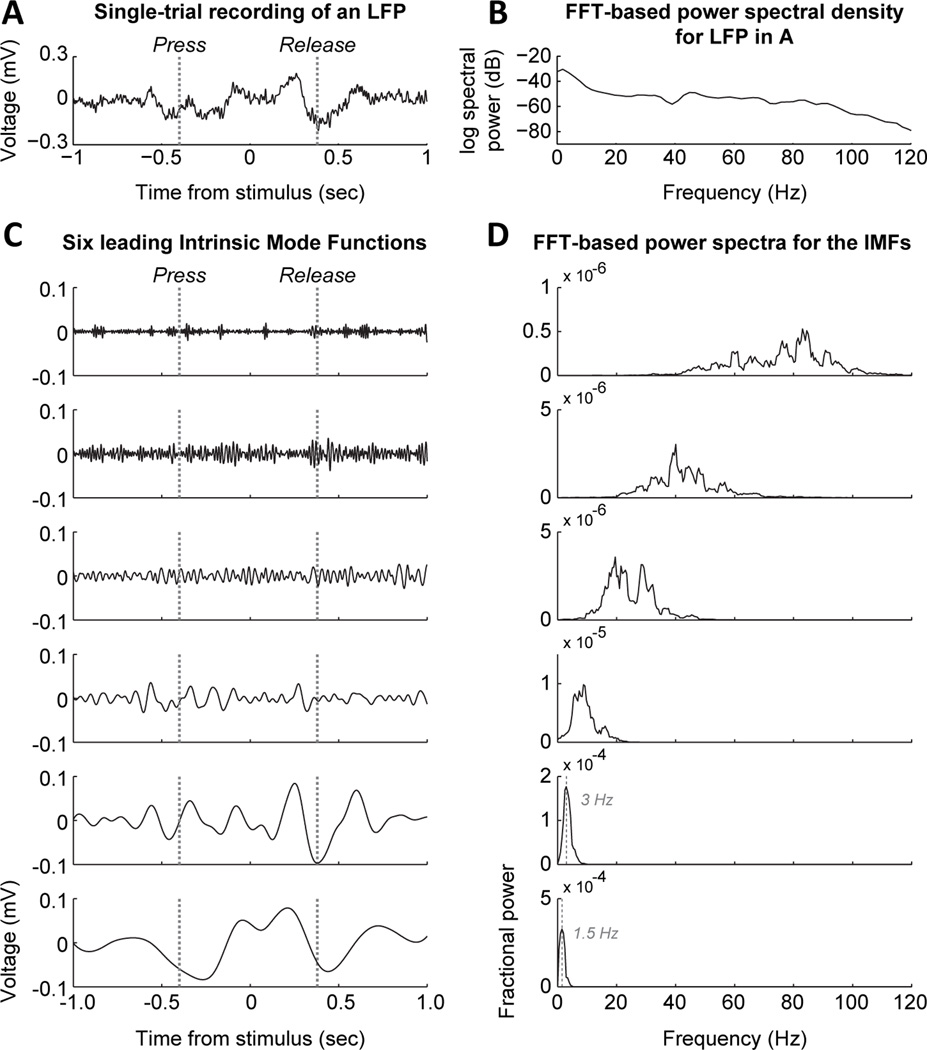
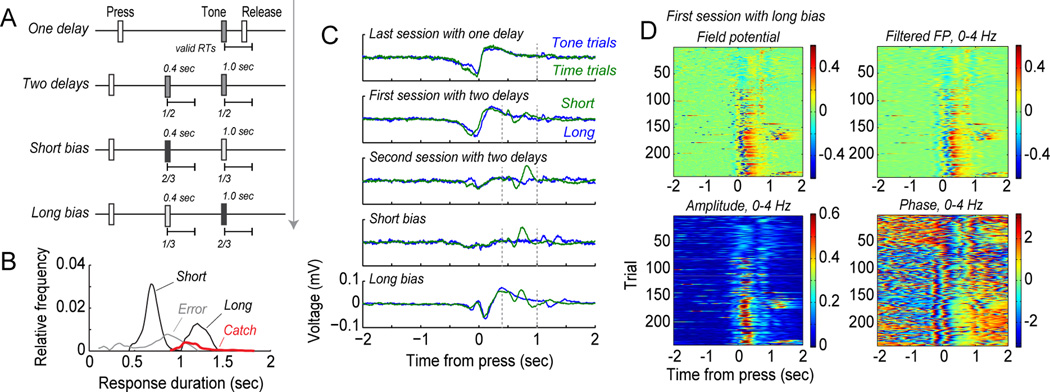
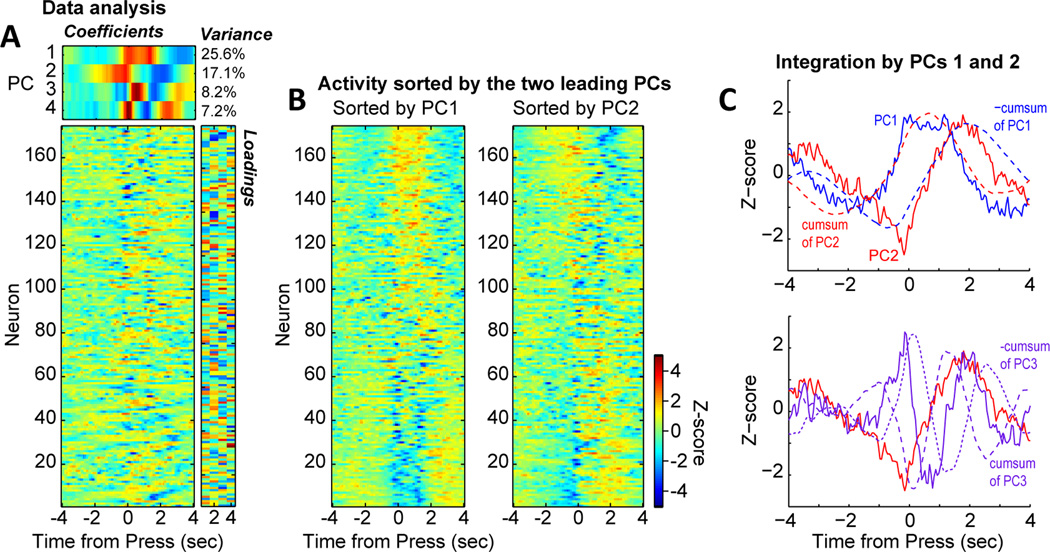
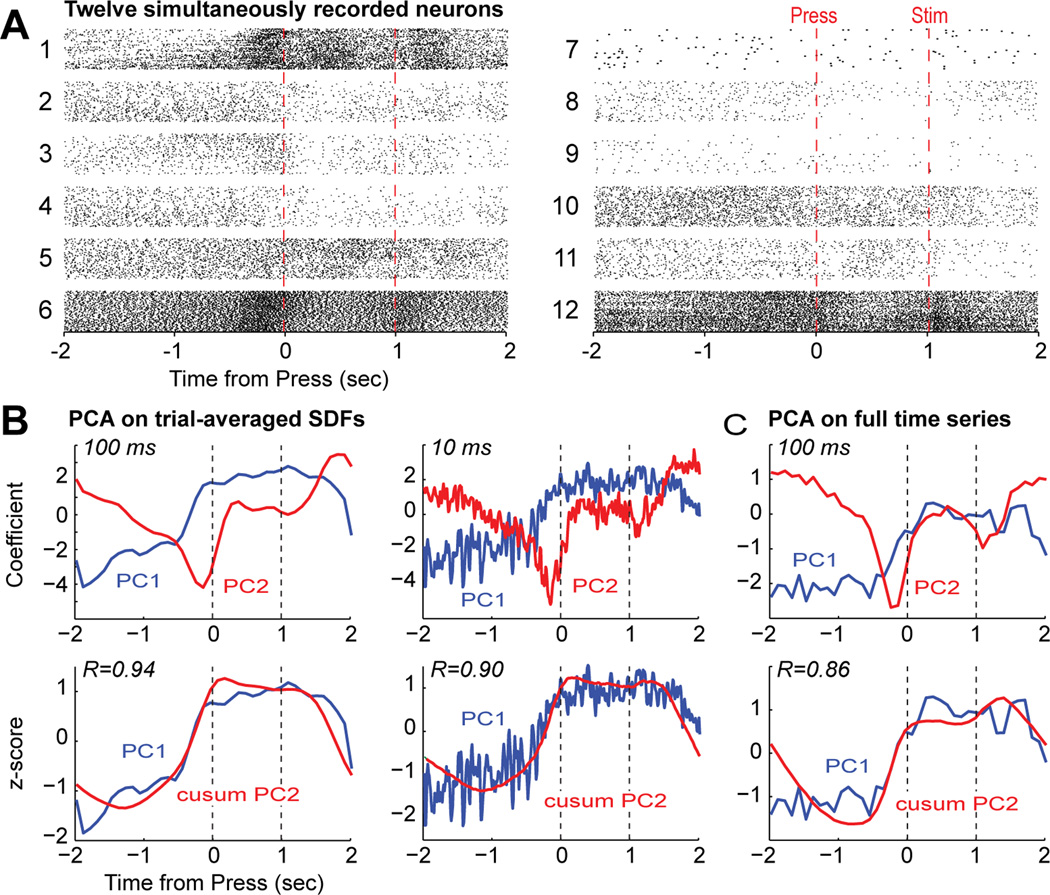
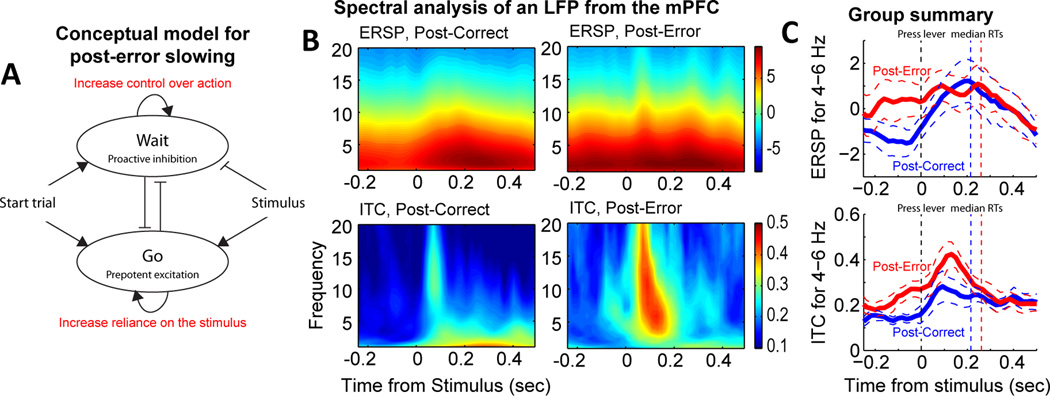
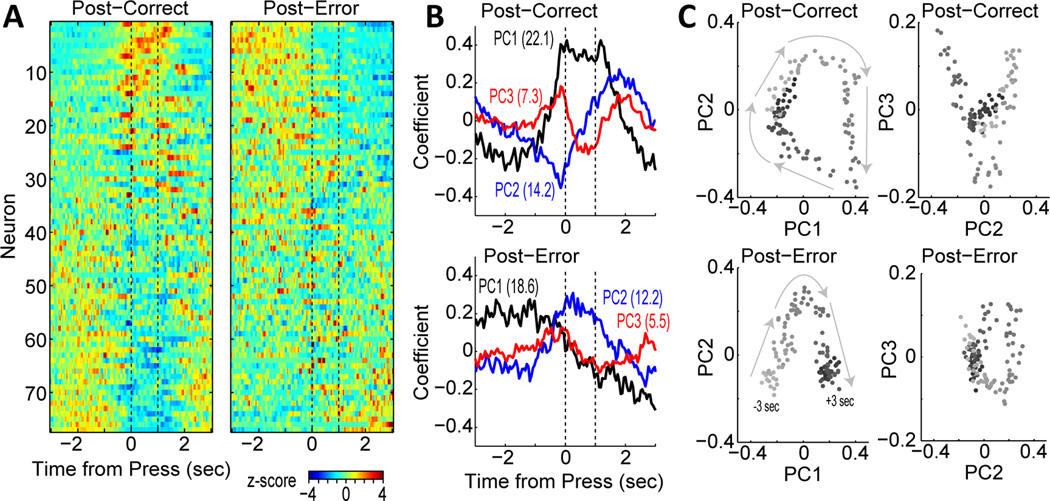
Studies in rats, monkeys and humans have established that the medial prefrontal cortex is crucial for the ability to exert adaptive control over behavior. Here, we review studies on the role of the rat medial prefrontal cortex in adaptive control, with a focus on simple reaction time tasks that can be easily used across species and have clinical relevance. The performance of these tasks is associated with neural activity in the medial prefrontal cortex that reflects stimulus detection, action timing, and outcome monitoring. We describe rhythmic neural activity that occurs when animals initiate a temporally extended action. Such rhythmic activity is coterminous with major changes in population spike activity. Testing animals over a series of sessions with varying pre-stimulus intervals showed that the signals adapt to the current temporal demands of the task. Disruptions of rhythmic neural activity occur on error trials (premature responding) and lead to a persistent encoding of the error and a subsequent change in behavioral performance (i.e. post-error slowing). Analysis of simultaneously recorded spike activity suggests that the presence of strong theta rhythms is coterminous with altered network dynamics, and might serve as a mechanism for adaptive control. Computational modeling suggests that these signals may enable learning from errors. Together, our findings contribute to an emerging literature and provide a new perspective on the neuronal mechanisms for the adaptive control of action.
Keywords: Anterior cingulate; Delta; Dynamics; Inhibition; Learning; Phase; Theta; Timing.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures
References
-
- Allen GV, Saper CB, Hurley KM, Cechetto DF. Organization of visceral and limbic connections in the insular cortex of the rat. J Comp Neurol. 1991;311:1–16. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
