

Kinetic characterization of the sole nonmuscle myosin-2 from the model organism Drosophila melanogaster
- PMID: 25636739
- PMCID: PMC4396609
- DOI: 10.1096/fj.14-266742
Kinetic characterization of the sole nonmuscle myosin-2 from the model organism Drosophila melanogaster
Abstract
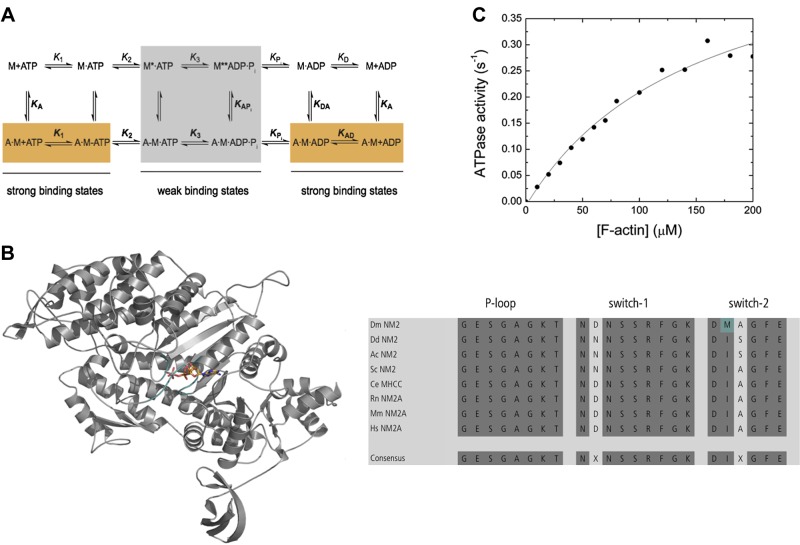
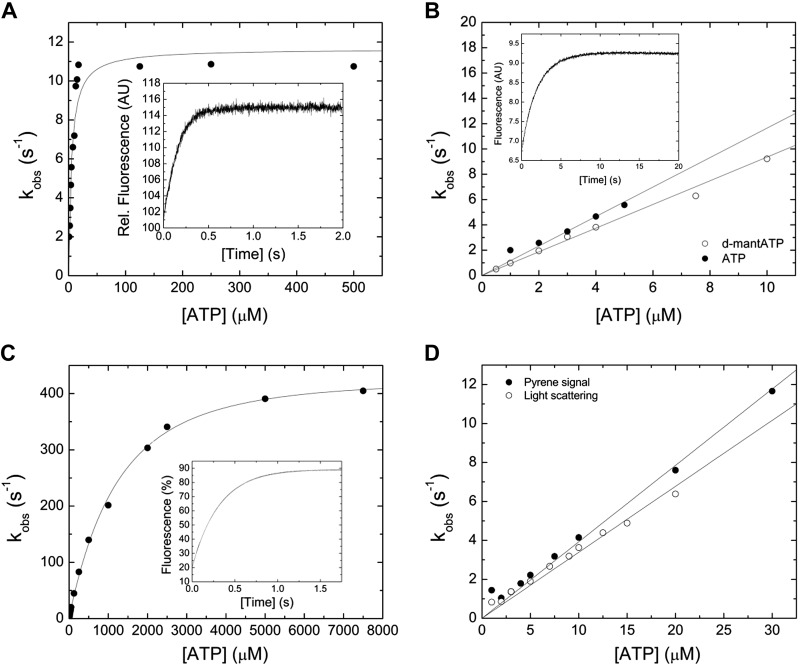
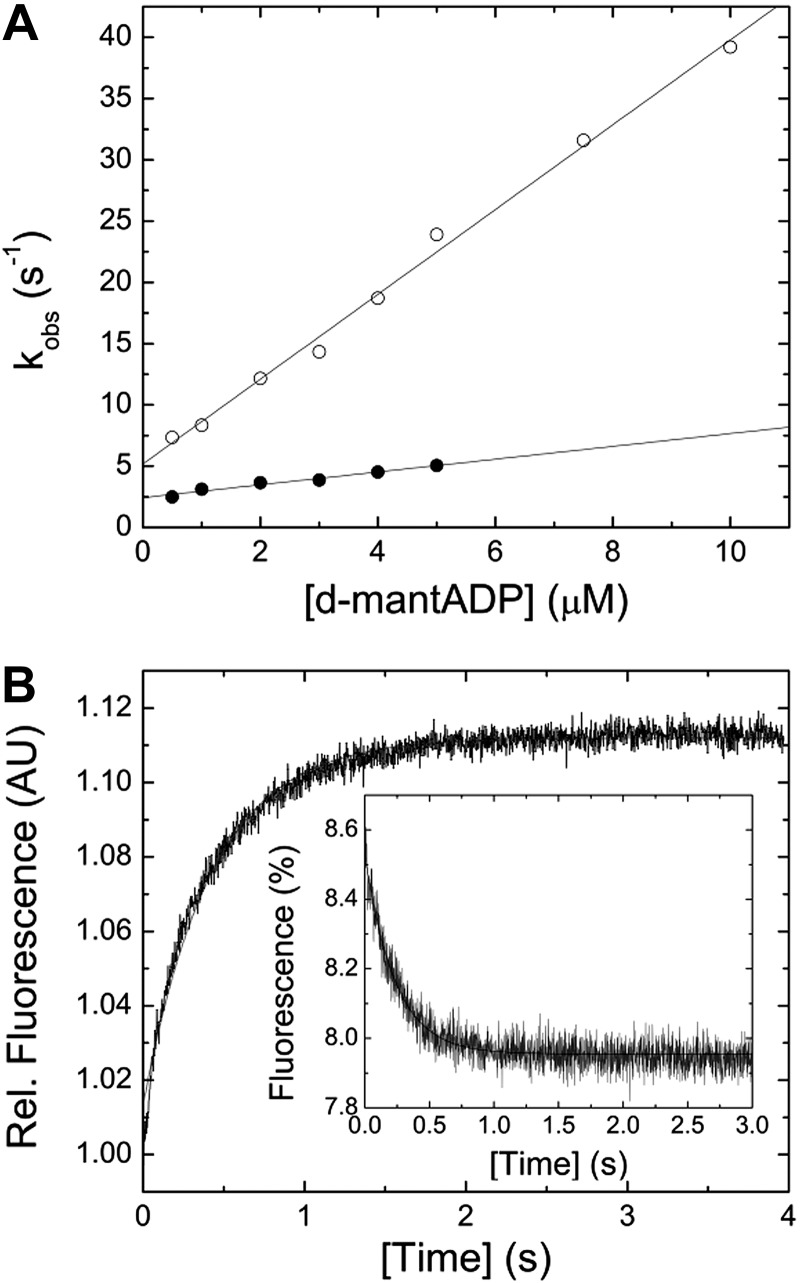
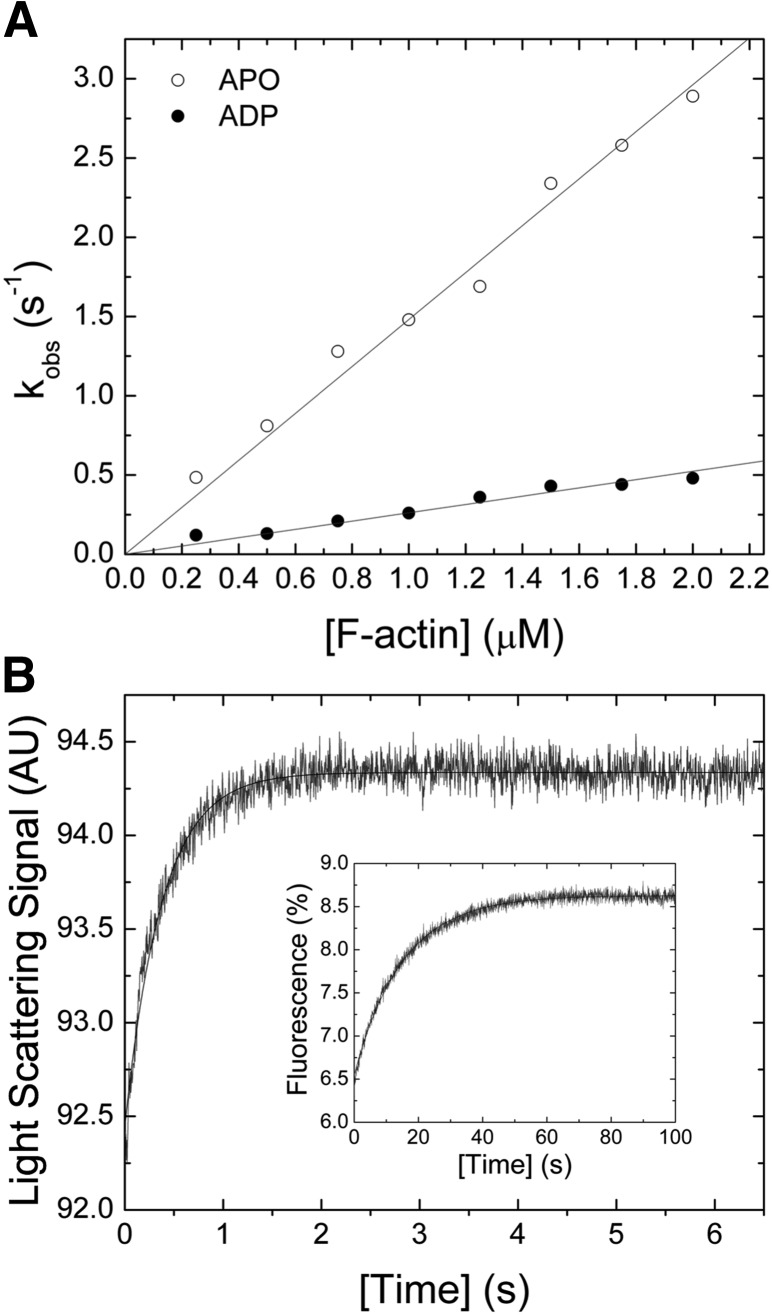
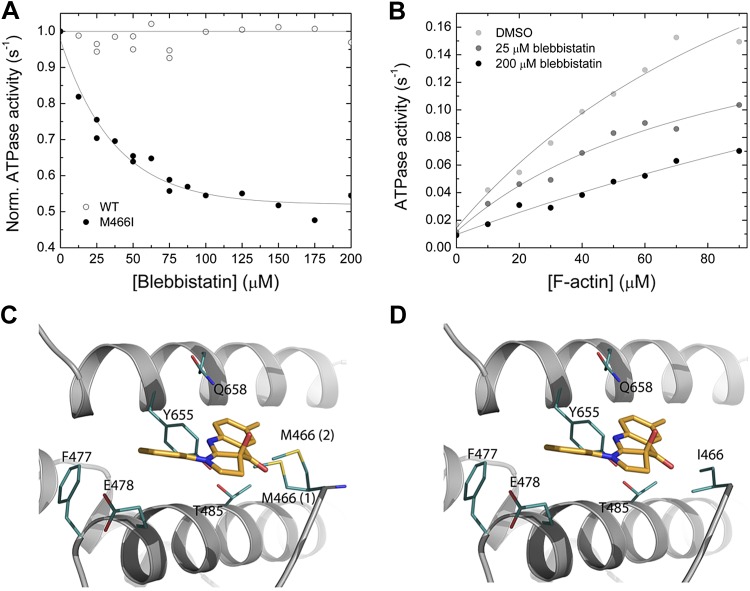
Nonmuscle myosin-2 is the primary enzyme complex powering contractility of the F-actin cytoskeleton in the model organism Drosophila. Despite myosin's essential function in fly development and homeostasis, its kinetic features remain elusive. The purpose of this in vitro study is a detailed steady-state and presteady-state kinetic characterization of the Drosophila nonmuscle myosin-2 motor domain. Kinetic features are a slow steady-state ATPase activity, high affinities for F-actin and ADP, and a low duty ratio. Comparative analysis of the overall enzymatic signatures across the nonmuscle myosin-2 complement from model organisms indicates that the Drosophila protein resembles nonmuscle myosin-2s from metazoa rather than protozoa, though modulatory aspects of myosin motor function are distinct. Drosophila nonmuscle myosin-2 is uniquely insensitive toward blebbistatin, a commonly used myosin-2 inhibitor. An in silico modeling approach together with kinetic studies indicate that the nonconsensus amino acid Met466 in the Drosophila nonmuscle myosin-2 active-site loop switch-2 acts as blebbistatin desensitizer. Introduction of the M466I mutation sensitized the protein for blebbistatin, resulting in a half-maximal inhibitory concentration of 36.3 ± 4.1 µM. Together, these data show that Drosophila nonmuscle myosin-2 is a bona fide molecular motor and establish an important link between switch-2 and blebbistatin sensitivity.
Keywords: actin; blebbistatin; cytoskeleton; rotamer; transient kinetics.
© FASEB.
Figures
References
-
- Young P. E., Richman A. M., Ketchum A. S., Kiehart D. P. (1993) Morphogenesis in Drosophila requires nonmuscle myosin heavy chain function. Genes Dev. 7, 29–41 - PubMed
-
- Sellers J. R. (2000) Myosins: a diverse superfamily. Biochim. Biophys. Acta 1496, 3–22 - PubMed
-
- Bloor J. W., Kiehart D. P. (2001) zipper Nonmuscle myosin-II functions downstream of PS2 integrin in Drosophila myogenesis and is necessary for myofibril formation. Dev. Biol. 239, 215–228 - PubMed
-
- Okumura T., Fujiwara H., Taniguchi K., Kuroda J., Nakazawa N., Nakamura M., Hatori R., Ishio A., Maeda R., Matsuno K. (2010) Left-right asymmetric morphogenesis of the anterior midgut depends on the activation of a non-muscle myosin II in Drosophila. Dev. Biol. 344, 693–706 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
