

Probing RNA-protein networks: biochemistry meets genomics
- PMID: 25636997
- PMCID: PMC4340748
- DOI: 10.1016/j.tibs.2015.01.003
Probing RNA-protein networks: biochemistry meets genomics
Abstract
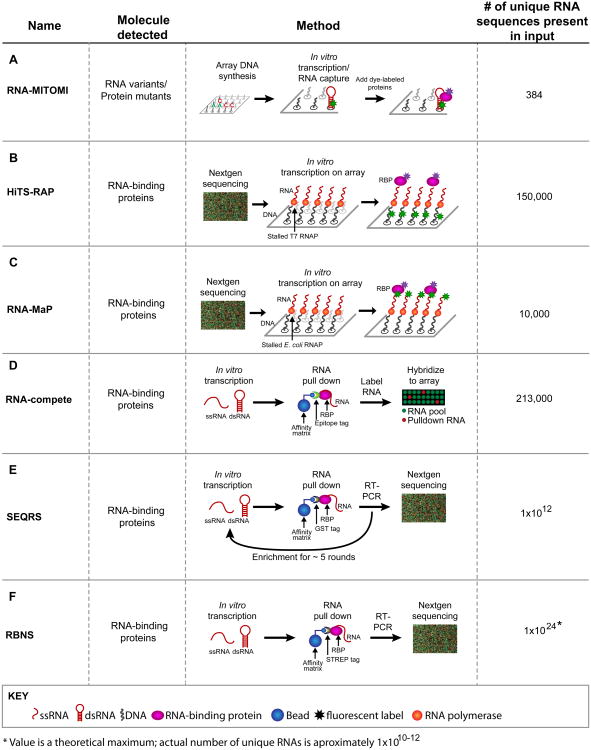
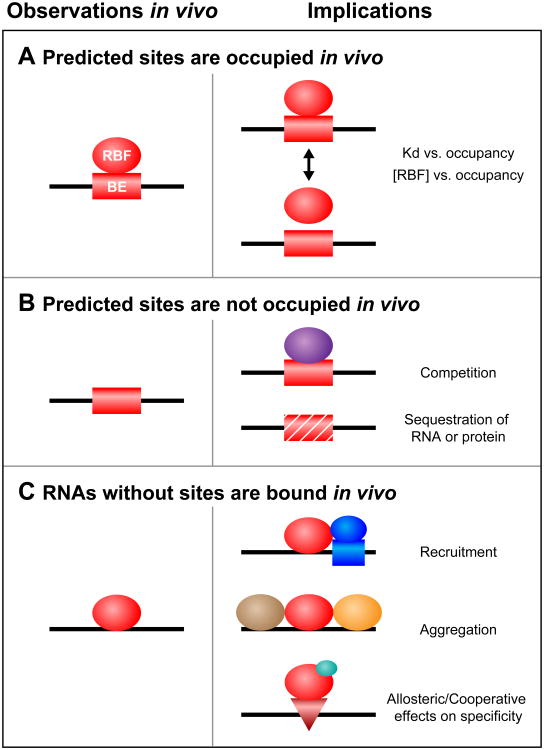
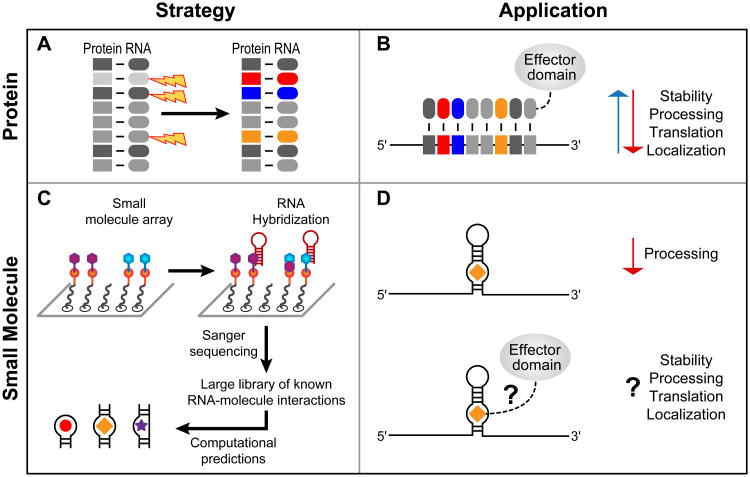
RNA-protein interactions are pervasive. The specificity of these interactions dictates which RNAs are controlled by what protein. Here we describe a class of revolutionary new methods that enable global views of RNA-binding specificity in vitro, for both single proteins and multiprotein complexes. These methods provide insight into central issues in RNA regulation in living cells, including understanding the balance between free and bound components, the basis for exclusion of binding sites, detection of binding events in the absence of discernible regulatory elements, and new approaches to targeting endogenous transcripts by design. Comparisons of in vitro and in vivo binding provide a foundation for comprehensive understanding of the biochemistry of protein-mediated RNA regulatory networks.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures
References
-
- Keene JD, et al. RIP-Chip: the isolation and identification of mRNAs, microRNAs and protein components of ribonucleoprotein complexes from cell extracts. Nat Protoc. 2006;1:302–307. - PubMed
-
- Guan L, Disney MD. Recent advances in developing small molecules targeting RNA. ACS Chem Biol. 2012;7:73–86. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
