

Induction of heme oxygenase-1 contributes to survival of Mycobacterium abscessus in human macrophages-like THP-1 cells
- PMID: 25638774
- PMCID: PMC4326180
- DOI: 10.1016/j.redox.2015.01.012
Induction of heme oxygenase-1 contributes to survival of Mycobacterium abscessus in human macrophages-like THP-1 cells
Abstract
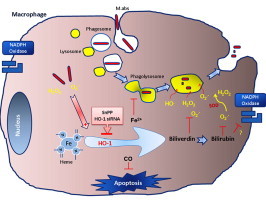
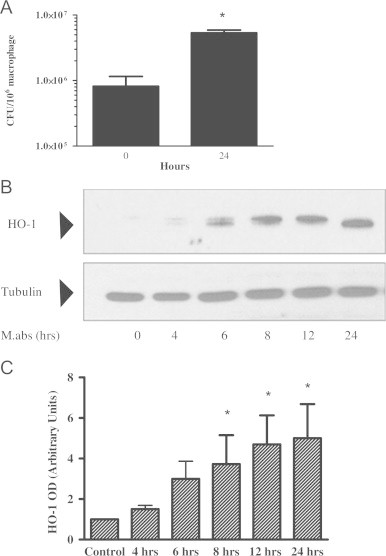
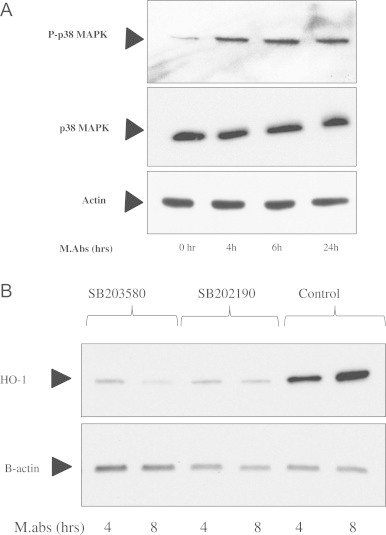
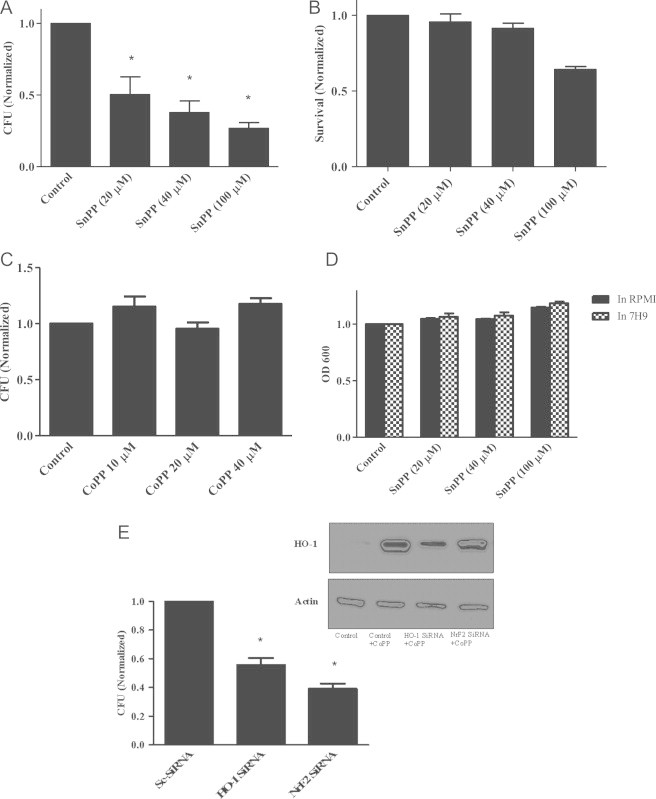
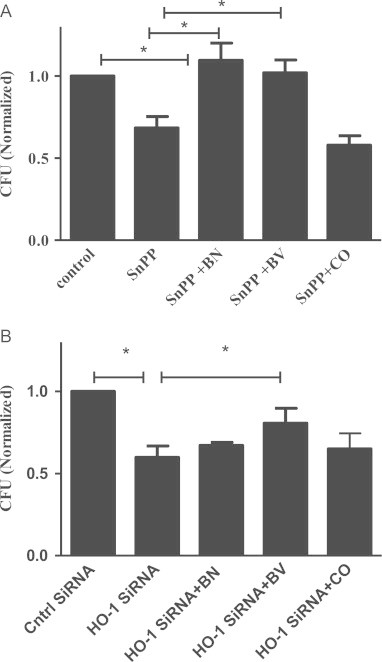
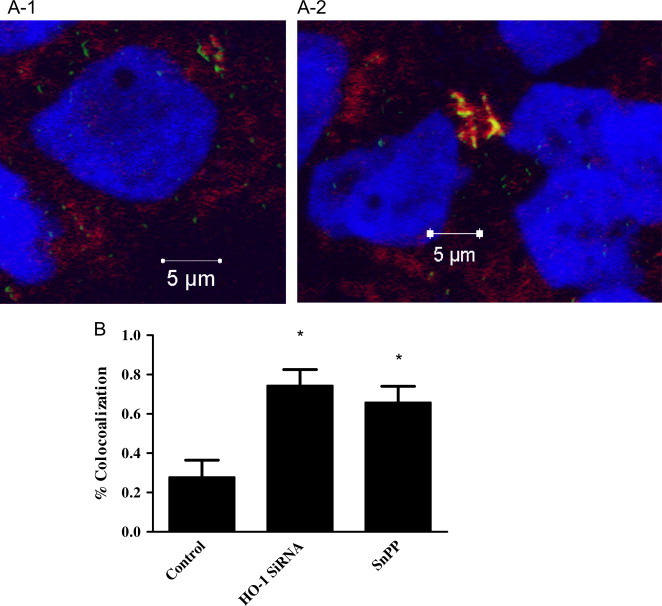
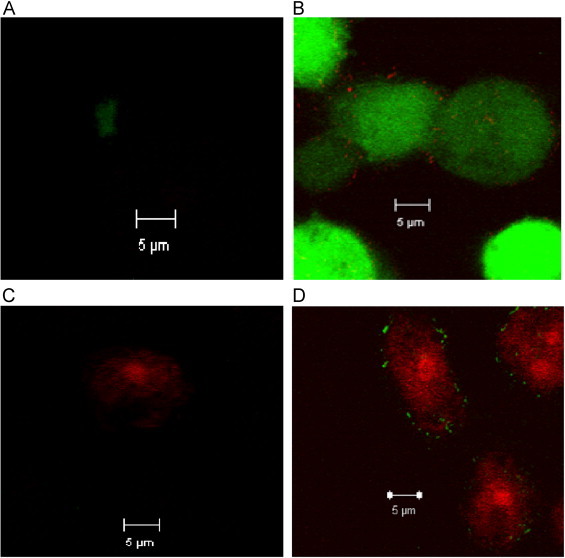
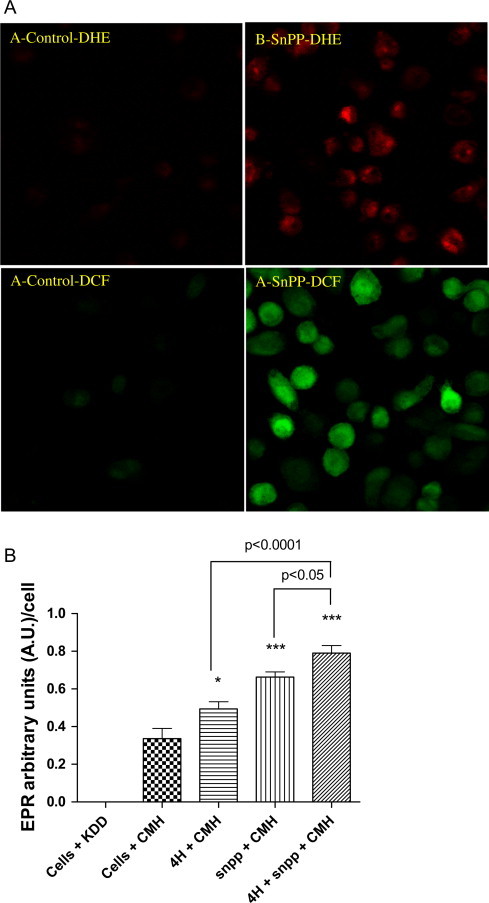
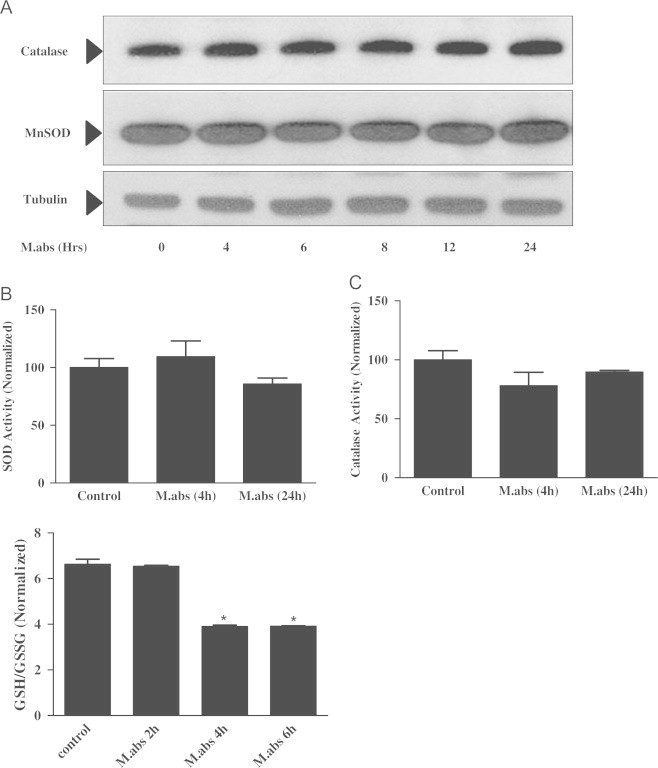
Mycobacterium abscessus (M.abs) is a rapidly growing mycobacterial species that infects macrophages, and is an important pathogen in patients with cystic fibrosis. We studied the early stages of M.abs infection of macrophages, with emphasis on the role of heme-oxygenase-1 (HO-1) in this infection. THP-1 cells were activated using TPA into macrophage-like cells and infected with M.abs for different time points. M.abs infection robustly induced HO-1 expression in the THP-1 cells. Production of HO-1 was p38 MAPK-dependent, as p38 inhibitors suppressed HO-1 induction. Pretreatment with HO-1 inhibitors tin-protoporphyrin (SnPP) significantly inhibited M.abs growth inside macrophages. Furthermore, inhibiting HO-1 using HO-1 siRNA or the HO-1 upstream signaling molecule; Nrf2 using Nrf2 siRNA resulted in similar inhibition of M.abs. In contrast, inducing HO-1 did not increase M.abs intracellular growth above control. Products of HO-1 metabolism of heme are bilirubin, biliverdin, carbon monoxide (CO) and iron. The addition of either bilirubin or biliverdin, but not CO, completely restored the SnPP inhibitory effect and partially that with HO-1 siRNA. To understand the mechanisms, we used Syto-62 labeled M.abs to infect macrophages. Interestingly, HO-1 inhibition promoted M.abs-containing phagosome fusion with lysosomes, which should enhance M.abs killing. M.abs infection enhanced THP-1 ROS production as demonstrated by increased DHE, DCF fluorescence, and EPR signal. HO-1 inhibition further increased ROS production in infected macrophages. Our results indicate that HO-1 induction is important for M.abs growth during the early stages of infection, and that the HO-1 products bilirubin and biliverdin, perhaps through modulation of intracellular ROS levels, may be involved.
Keywords: HO-1; Mycobacterium abscessus; Oxidative stress; THP-1 cells.
Copyright © 2015 The Authors. Published by Elsevier B.V. All rights reserved.
Figures
References
-
- Román A., Ussetti P., Solé A., Zurbano F., Borro J.M., Vaquero J.M., de Pablo A., Morales P., Blanco M., Bravo C., Cifrian J., de la Torre M., Gámez P., Laporta R., Monforte V., Mons R., Salvatierra A., Santos F., Solé J., Varela A. Guidelines for the selection of lung transplantation candidates. Archivos de BronconeumologÃa. 2011;47(6):303–309. doi: 10.1016/j.arbres.2011.03.007. English Version. - DOI - PubMed
-
- Bryant J.M., Grogono D.M., Greaves D., Foweraker J., Roddick I., Inns T., Reacher M., Haworth C.S., Curran M.D., Harris S.R., Peacock S.J., Parkhill J., Floto R.A. Whole-genome sequencing to identify transmission of Mycobacterium abscessus between patients with cystic fibrosis: a retrospective cohort study. Lancet. 2013;381(9877):1551–1560. doi: 10.1016/S0140-6736(13)60632-7. 23541540 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
