

Amino acid substitutions in the non-structural proteins 4A or 4B modulate the induction of autophagy in West Nile virus infected cells independently of the activation of the unfolded protein response
- PMID: 25642225
- PMCID: PMC4295549
- DOI: 10.3389/fmicb.2014.00797
Amino acid substitutions in the non-structural proteins 4A or 4B modulate the induction of autophagy in West Nile virus infected cells independently of the activation of the unfolded protein response
Abstract
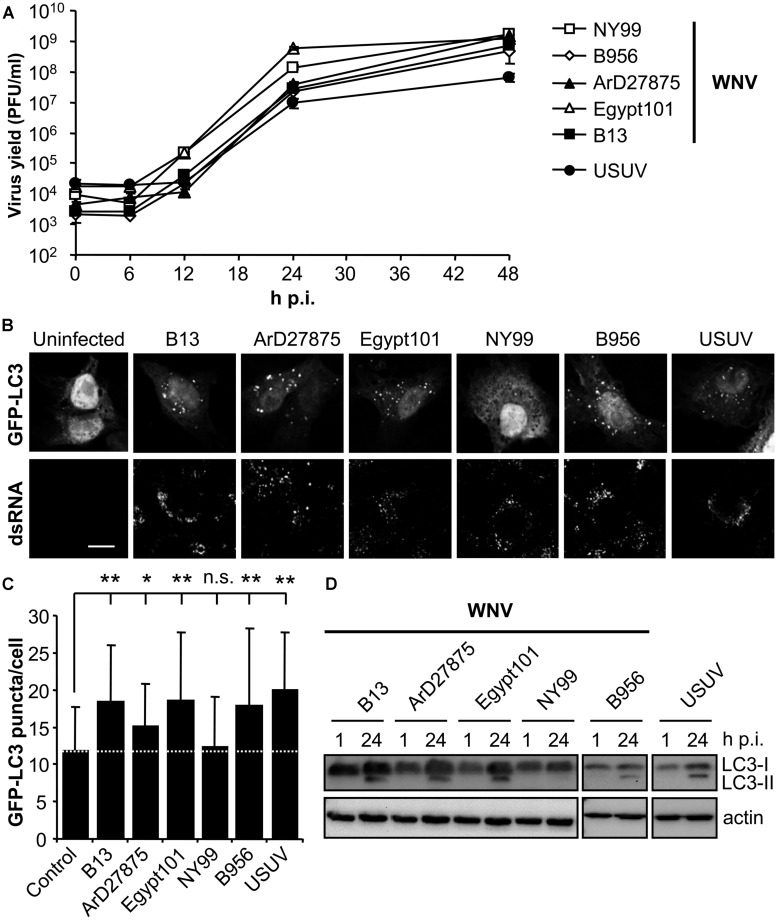
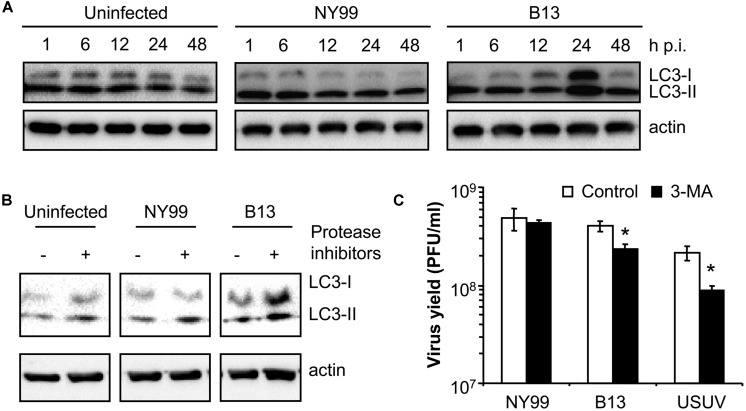
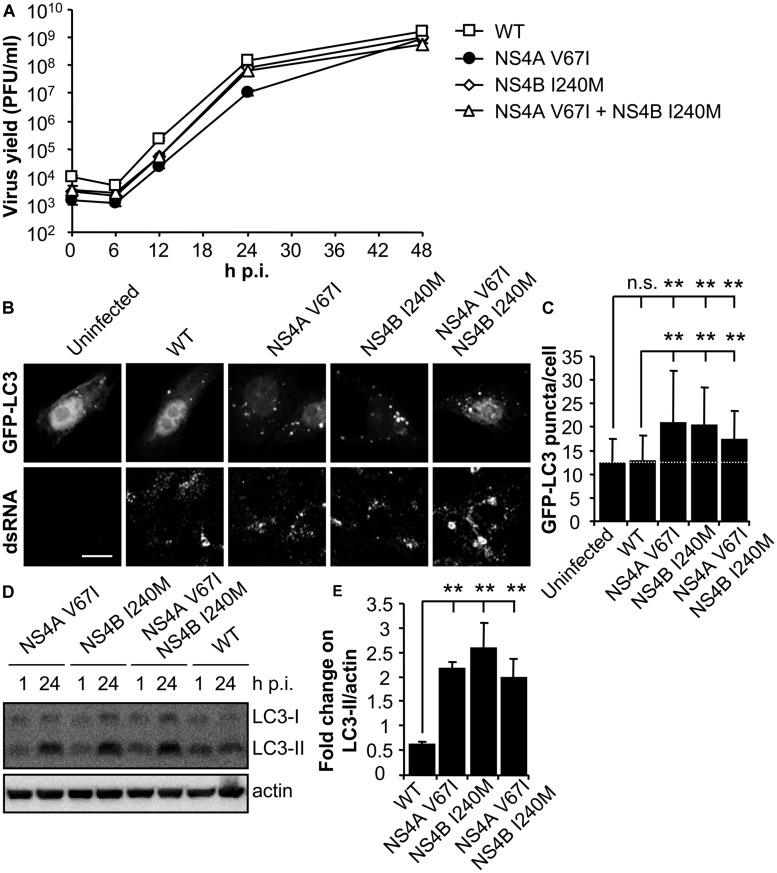
West Nile virus (WNV) is a neurotropic mosquito-borne flavivirus responsible for outbreaks of meningitis and encephalitis. Whereas the activation of autophagy in cells infected with other flaviviruses is well known, the interaction of WNV with the autophagic pathway still remains unclear and there are reports describing opposite findings obtained even analyzing the same viral strain. To clarify this controversy, we first analyzed the induction of autophagic features in cells infected with a panel of WNV strains. WNV was determined to induce autophagy in a strain dependent manner. We observed that all WNV strains or isolates analyzed, except for the WNV NY99 used, upregulated the autophagic pathway in infected cells. Interestingly, a variant derived from this WNV NY99 isolated from a persistently infected mouse increased LC3 modification and aggregation. Genome sequencing of this variant revealed only two non-synonymous nucleotide substitutions when compared to parental NY99 strain. These nucleotide substitutions introduced one amino acid replacement in NS4A and other in NS4B. Using genetically engineered viruses we showed that introduction of only one of these replacements was sufficient to upregulate the autophagic pathway. Thus, in this work we have shown that naturally occurring point mutations in the viral non-structural proteins NS4A and NS4B confer WNV with the ability to induce the hallmarks of autophagy such as LC3 modification and aggregation. Even more, the differences on the induction of an autophagic response observed among WNV variants in infected cells did not correlate with alterations on the activation of the unfolded protein response (UPR), suggesting an uncoupling of UPR and autophagy during flavivirus infection. The findings here reported could help to improve the knowledge of the cellular processes involved on flavivirus-host cell interactions and contribute to the design of effective strategies to combat these pathogens.
Keywords: LC3; West Nile virus (WNV); autophagy; host cells; replication; unfolded protein response.
Figures
Similar articles
-
Reconciling West Nile virus with the autophagic pathway.Autophagy. 2015;11(5):861-4. doi: 10.1080/15548627.2015.1037062. Autophagy. 2015. PMID: 25946067 Free PMC article.
-
Induction of virus-specific effector immune cell response limits virus replication and severe disease in mice infected with non-lethal West Nile virus Eg101 strain.J Neuroinflammation. 2015 Sep 22;12:178. doi: 10.1186/s12974-015-0400-y. J Neuroinflammation. 2015. PMID: 26392176 Free PMC article.
-
Autophagy inhibits viral genome replication and gene expression stages in West Nile virus infection.Virus Res. 2014 Oct 13;191:83-91. doi: 10.1016/j.virusres.2014.07.016. Epub 2014 Aug 1. Virus Res. 2014. PMID: 25091564
-
Stress responses in flavivirus-infected cells: activation of unfolded protein response and autophagy.Front Microbiol. 2014 Jun 3;5:266. doi: 10.3389/fmicb.2014.00266. eCollection 2014. Front Microbiol. 2014. PMID: 24917859 Free PMC article. Review.
-
Virulence determinants of West Nile virus: how can these be used for vaccine design?Future Virol. 2017 May;12(5):283-295. doi: 10.2217/fvl-2016-0141. Epub 2017 Apr 28. Future Virol. 2017. PMID: 28919920 Free PMC article. Review.
Cited by
-
Role of autophagy during the replication and pathogenesis of common mosquito-borne flavi- and alphaviruses.Open Biol. 2019 Mar 29;9(3):190009. doi: 10.1098/rsob.190009. Open Biol. 2019. PMID: 30862253 Free PMC article. Review.
-
Divergent Mutational Landscapes of Consensus and Minority Genotypes of West Nile Virus Demonstrate Host and Gene-Specific Evolutionary Pressures.Genes (Basel). 2020 Oct 30;11(11):1299. doi: 10.3390/genes11111299. Genes (Basel). 2020. PMID: 33143358 Free PMC article.
-
N-Desmethylclozapine, Fluoxetine, and Salmeterol Inhibit Postentry Stages of the Dengue Virus Life Cycle.Antimicrob Agents Chemother. 2016 Oct 21;60(11):6709-6718. doi: 10.1128/AAC.01367-16. Print 2016 Nov. Antimicrob Agents Chemother. 2016. PMID: 27572397 Free PMC article.
-
Host Subcellular Organelles: Targets of Viral Manipulation.Int J Mol Sci. 2024 Jan 29;25(3):1638. doi: 10.3390/ijms25031638. Int J Mol Sci. 2024. PMID: 38338917 Free PMC article. Review.
-
Virus, Exosome, and MicroRNA: New Insights into Autophagy.Adv Exp Med Biol. 2022;1401:97-162. doi: 10.1007/5584_2022_715. Adv Exp Med Biol. 2022. PMID: 35781219
References
-
- Bakonyi T., Gould E. A., Kolodziejek J., Weissenbock H., Nowotny N. (2004). Complete genome analysis and molecular characterization of Usutu virus that emerged in Austria in 2001: comparison with the South African strain SAAR-1776 and other flaviviruses. Virology 328 301–310 10.1016/j.virol.2004.08.005 - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources