

Alpha-enolase is upregulated on the cell surface and responds to plasminogen activation in mice expressing a ∆133p53α mimic
- PMID: 25643152
- PMCID: PMC4313950
- DOI: 10.1371/journal.pone.0116270
Alpha-enolase is upregulated on the cell surface and responds to plasminogen activation in mice expressing a ∆133p53α mimic
Abstract
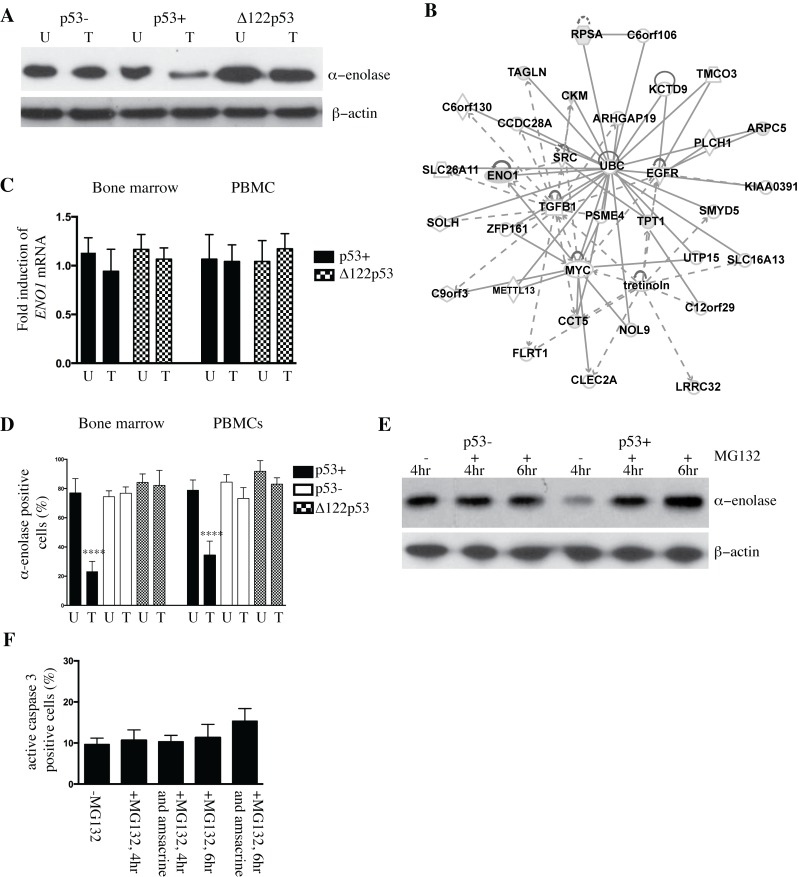
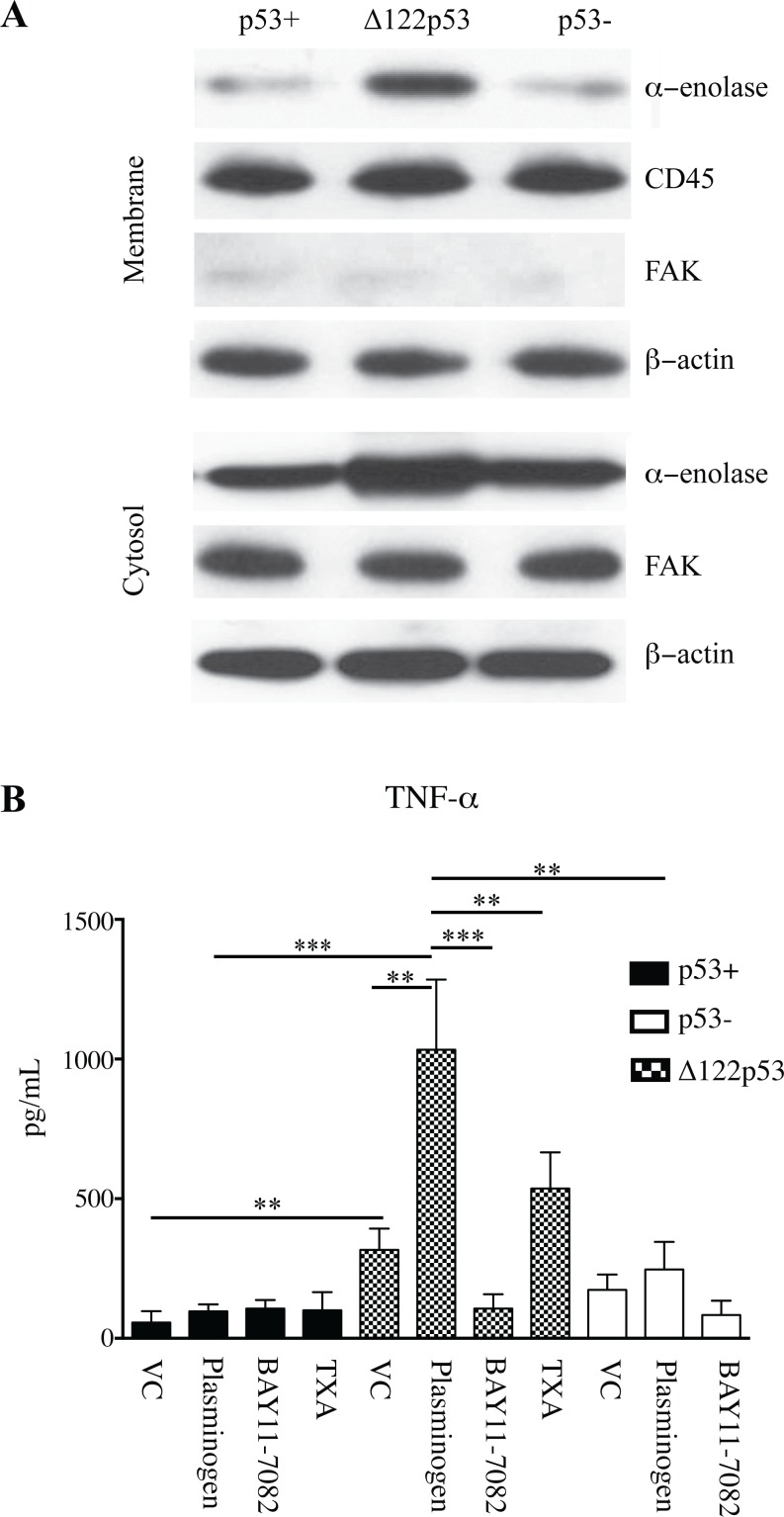
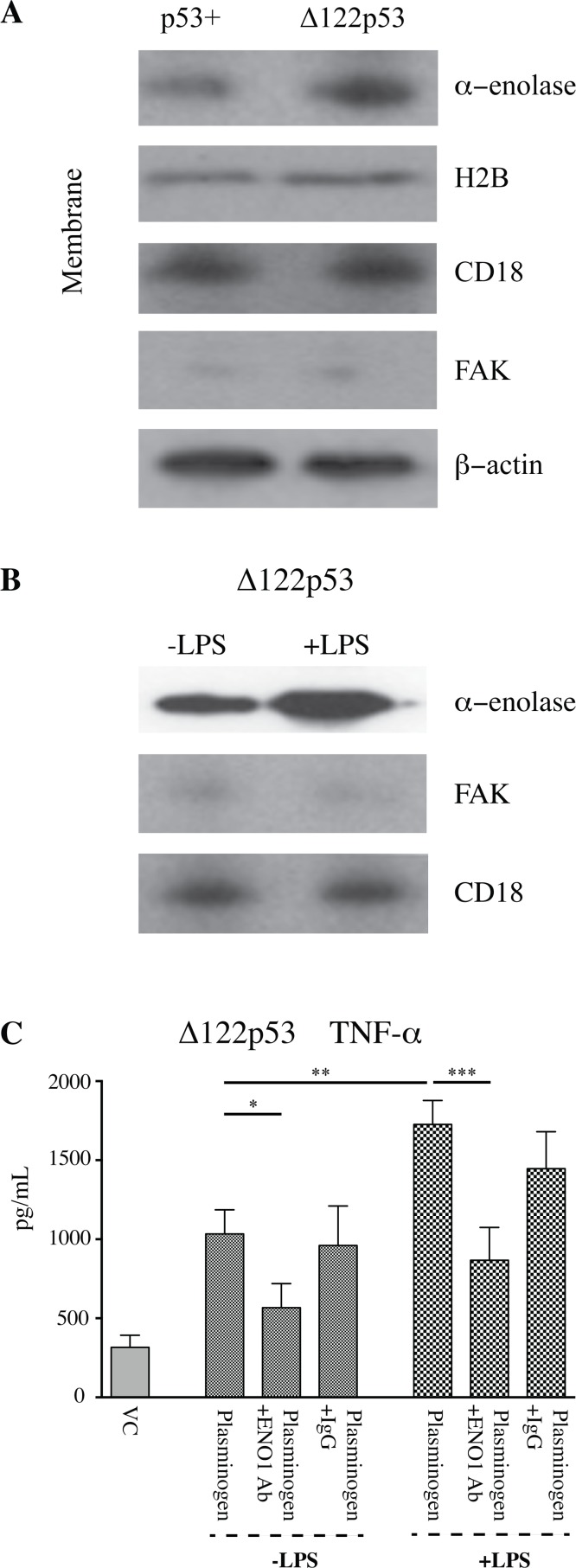
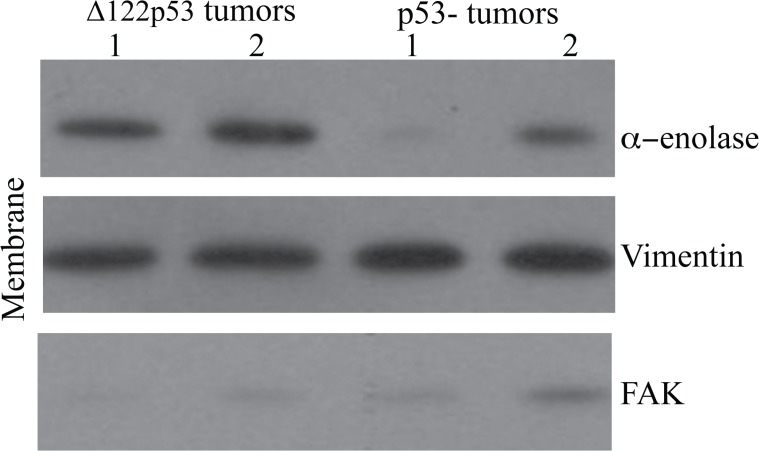
The p53 protein is a master regulator of the stress response. It acts as a tumor suppressor by inducing transcriptional activation of p53 target genes, with roles in apoptosis, cell cycle arrest and metabolism. The discovery of at least 12 isoforms of p53, some of which have tumor-promoting properties, has opened new avenues of research. Our previous work studied tumor phenotypes in four mouse models with different p53 backgrounds: wild-type p53, p53 null, mutant p53 lacking the proline domain (mΔpro), and a mimic for the human Δ133p53α p53 isoform (Δ122p53). To identify the major proteins affected by p53 function early in the response to DNA damage, the current study investigated the entire proteome of bone marrow, thymus, and lung in the four p53 models. Protein extracts from untreated controls and those treated with amsacrine were analyzed using two-dimensional fluorescence difference gel electrophoresis. In the bone marrow, reactive proteins were universally decreased by wild-type p53, including α-enolase. Further analysis of α-enolase in the p53 models revealed that it was instead increased in Δ122p53 hematopoietic and tumor cell cytosol and on the cell surface. Alpha-enolase on the surface of Δ122p53 cells acted as a plasminogen receptor, with tumor necrosis factor alpha induced upon plasminogen stimulation. Taken together, these data identified new proteins associated with p53 function. One of these proteins, α-enolase, is regulated differently by wild-type p53 and Δ122p53 cells, with reduced abundance as part of a wild-type p53 response and increased abundance with Δ122p53 function. Increased cell surface α-enolase on Δ122p53 cells provides a possible explanation for the model's pro-inflammatory features and suggests that p53 isoforms may direct an inflammatory response by increasing the amount of α-enolase on the cell surface.
Conflict of interest statement
Figures
References
-
- Oren M (2003) Decision making by p53: life, death and cancer. Cell Death Differ 10: 431–442. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
